Table of Contents
The Structure and Mechanism of an Active, Lipid-Linked Flippase
This Wiki page is adapted from a 2015 Nature publication by Perez et al. entitled “Structure and Mechanism of an Active, Lipid-Linked Oligosaccharide Flippase”. Read the full article here: https://www.nature.com/articles/nature14953
Introduction
Several cellular processes in bacteria require the transport of molecules from the inside to the outside of the cell for the synthesis of extracellular structures or addition to other molecules. One such process is N-glycosylation, which requires oligosaccharides to be transported from the cytoplasm to periplasm and subsequently added to membrane-bound proteins (Perez et al., 2015). These oligosaccharides are linked to a large lipid molecule embedded in the cellular membrane. However, flipping of these membrane-embedded lipids with polar head groups is slow and energetically unfavorable (Perez et al., 2015). Thus, these reactions must be catalyzed by flippases. In Campylobacter jejuni, the flipping of lipid linked oligosaccharides during protein N-glycosylation is catalyzed by the flippase PglK; however, its mechanism has yet to be elucidated. Addressing this gap in knowledge, a 2015 Nature paper published by Perez et al. outlines the structure and mechanism of the active, lipid-linked oligosaccharide flippase PglK. The following Wiki page will further outline the role of flippases, their essentiality in N-glycosylation, and the specific role of PglK in C. jejuni.
About the Authors
Plagued by the lack of knowledge surrounding how PglK is able to flip oligosaccharides, scientists from an array of Suisse institutions collaborated with one common goal: elucidating the structure and mechanism of PglK.
Dr. Camilo Perez
 Lead researcher, Dr. Camilo Perez, is the Head of a Research Group at the Biozentrum Center for Molecular Life Sciences at the University of Basel in Switzerland (Biozentrum, 2020). His group aims to understand the molecular mechanism of membrane proteins involved in the assembly of the bacterial cell wall (Biozentrum, 2020). Transporters, such as flippases, are one type of membrane protein involved in these processes. As such, Dr. Perez’s work in elucidating the structure and function of membrane proteins led to his interest in the transport protein PglK.
Lead researcher, Dr. Camilo Perez, is the Head of a Research Group at the Biozentrum Center for Molecular Life Sciences at the University of Basel in Switzerland (Biozentrum, 2020). His group aims to understand the molecular mechanism of membrane proteins involved in the assembly of the bacterial cell wall (Biozentrum, 2020). Transporters, such as flippases, are one type of membrane protein involved in these processes. As such, Dr. Perez’s work in elucidating the structure and function of membrane proteins led to his interest in the transport protein PglK.
Dr. Kaspar P. Locher
 Dr. Kaspar P. Locher is a researcher in the Department of Biology at ETH Zurich. His lab studies the structure and mechanism of membrane proteins involved in active transport, leading to his interest in elucidating the role of PglK (ETH Zurich, 2023).
Dr. Kaspar P. Locher is a researcher in the Department of Biology at ETH Zurich. His lab studies the structure and mechanism of membrane proteins involved in active transport, leading to his interest in elucidating the role of PglK (ETH Zurich, 2023).
Dr. Sabina Gerber
 Dr. Sabina Gerber is head of the Centre for Biochemistry and Bioanalytics at the ZHAW University of Applied Sciences in Switzerland. Her assistance with this project was a natural extension of her previous work on the structural determination of the oligosaccharyltransferase PglB which is also involved in bacterial N-glycosylation.
Dr. Sabina Gerber is head of the Centre for Biochemistry and Bioanalytics at the ZHAW University of Applied Sciences in Switzerland. Her assistance with this project was a natural extension of her previous work on the structural determination of the oligosaccharyltransferase PglB which is also involved in bacterial N-glycosylation.
Dr. Jean-Louis Reymond
 Dr. Jean-Louis Reymond is a Researcher in the Department of Chemistry, Biochemistry and Pharmaceutical Sciences at the University of Bern in Switzerland. His lab uses computational tools to analyze the existing structures and properties of molecules, to develop new drugs, and to study molecule synthesis. He designed and synthesized fluorescent peptides and lipid-linked oligosaccharide model substrates for use in this study (Reymond Research Group, 2023).
Dr. Jean-Louis Reymond is a Researcher in the Department of Chemistry, Biochemistry and Pharmaceutical Sciences at the University of Bern in Switzerland. His lab uses computational tools to analyze the existing structures and properties of molecules, to develop new drugs, and to study molecule synthesis. He designed and synthesized fluorescent peptides and lipid-linked oligosaccharide model substrates for use in this study (Reymond Research Group, 2023).
Campylobacter jejuni
The model organism in this paper is the gram-negative bacterium, Campylobacter jejuni. This bacterium has been the paradigm organism to study bacterial N-glycosylation pathways since the discovery of these pathways in 2002. C. jejuni has a well-characterized N-glycosylation system that modifies over 60 periplasmic and membrane-bound proteins (Alemka et al., 2013). Protein glycosylation is essential in C. jejuni because it mediates the organism’s immunogenicity and pathogenicity. Specifically, C. jejuni is a gut pathogen and the causative agent of human bacterial gastroenteritis (Alemka et al., 2013). Research by Alemka et al. (2013) demonstrated that N-glycosylation of surface proteins enhances C. jejuni’s fitness by protecting bacterial proteins from cleavage by gut proteases. Additionally, glycosylation facilitates colonization and invasion of intestinal epithelial cells (Alemka et al., 2013). As flippases play an essential role in glycosylating these surface proteins, a clearer understanding of the flippase PglK’s structure and mechanism can provide crucial insight into the pathogenesis of C. jejuni.
Flippases
What is a Flippase and Why are they Important?

Figures 1 & 2: Flippase enzyme catalyzing the translocation of a phospholipid across the membrane. On the left, the general function of a flippase is shown, as it flips a phospholipid from one leaflet to the other. On the right, the catalysis of lipid translocation is shown. The top image depicts the process as slow and energetically costly in the absence of a flippase. The bottom image depicts the process occuring faster when a flippase is present.
Flippases are membrane-bound enzymes responsible for transporting phospholipid molecules between the extracellular leaflet and the cytosolic leaflet of the cell membrane (Figure 1). Although lipid components are able to diffuse through the membrane, their polar head groups cannot easily pass through the hydrophobic interior (Hankins et al., 2015). Thus, transporting these molecules is slow and energetically unfavorable. To overcome this energy barrier, flippases act as a “pump” to catalyze the movement of lipids between leaflets, essentially flipping the compound (Figure 2). When this function was discovered by Mark Brescher in the early 1970s, he coined the term “flippase” to describe the enzyme responsible (Hankins et al., 2015).
Flippases have the primary responsibility of maintaining an asymmetric distribution of lipids across the bilayer (Figure 3) (Graham, 2021). They concentrate phosphatidylserine (PS) and phosphatidylethanolamine (PE) to the cytosolic leaflet of the membrane by removing them from the extracellular leaflet. Phosphatidylcholine (PC) and sphingomyelin (SM) are enriched in the opposite manner, as they are concentrated in the extracellular leaflet (Clarke et al., 2020). Flippases are essential for maintaining this non-equilibrium state by flipping PS, PE, PC and SM from one leaflet to the other. These phospholipids have differing net charges at a physiological pH. Their translocation generates a large contrast in charge between the two leaflets, creating asymmetry in the membrane (University of Singapore, 2018).

Figures 3 & 4: Flippase maintaining lipid asymmetry & providing molecules for extracellular structures. On the left, a flippase is shown maintaining lipid asymmetry by translocating certain phospholipids to either side of the membrane. Namely, PS is enriched in the cytosolic leaflet. On the right, a flippase is shown flipping a lipid II molecule across the membrane to be used in the structure of the peptidoglycan.
This lipid asymmetry is vital in numerous eukaryotic physiological processes, namely signal transduction. PS binds and regulates various signaling proteins, such as protein kinase C. Also, PS translocation is involved with the recognition and elimination of apoptotic cells. PS exposure on the outer leaflet indicates that the integrity of the membrane has been lost, providing a signal to surrounding macrophages to engulf the cell (Clarke et al., 2018). Therefore, lipid asymmetry is crucial in the maintenance of cell homeostasis. Lipid asymmetry and translocation further influence membrane curvature, vesicle formation, and the triggering of membrane associated events. Flippases contribute to these important cellular processes through their role of maintaining asymmetry across the lipid bilayer (Devaux et al., 2018).
Another reason that flippases are essential to cells is because other molecules, like sugars, can be linked to these lipids and then flipped across the cell membrane (Hankins et al., 2015). This often occurs in bacterial cells during the synthesis of extracellular structures, like lipopolysaccharide (Hankins et al., 2015). An additional example occurs in peptidoglycan assembly, as the lipid II flippase, MurJ, flips lipid II molecules across the membrane where they are subsequently added to the growing cell wall (Figure 4). Moreover, in bacterial N-glycosylation, a flippase is required to transport lipid-linked oligosaccharides (LLO) across the membrane for attachment to proteins (Hankins et al., 2015). This is the process that is detailed throughout the paper by Perez et al. (2015).
Types of Flippases

Figure 5: Different types of flippases. A scramblase (green) is shown on the left and can translocate phospholipids bidirectionally. It does not rely on ATP to catalyze this reaction. A P4-ATPase (blue) is shown in the middle, and translocates the lipids from the outer leaflet to the inner leaflet by utilizing ATP. In contrast, the ABC transporter (pink) on the right uses ATP to flip the lipids from the inner leaflet to the outer leaflet.
Passive Transporters
Passive–or energy independent transporters–include scramblase proteins that focus on randomizing the 3-dimensional orientation of lipids across the bilayer, as well as proton/sodium driven secondary antiporters (Perez et al., 2015). These ATP-independent proteins can facilitate bidirectional transport of lipids across the membrane (Sharom, 2011).
Active Transporters
Active transporters are those that specifically rely on the use of ATP energy to catalyze the active translocation of lipids across the cell membrane (Perez et al., 2015). These ATP-dependent transporters include P4-ATPases and ATP-binding cassette (ABC) transporters (Perez et al., 2015).
P4-ATPases catalyze the movement of phospholipids from the extracellular leaflet to the cytosolic leaflet. They differ in their substrate specificities and mediate transport of a broader range of lipid substrates, including lysophospholipids and synthetic alkylphospholipids (Lopez-Marques et al., 2014). Based on sequence similarity, the P-type ATPase family is divided into five subfamilies (P1–P5) with different transport specificities (Lopez-Marques et al., 2014).
ABC transporters catalyze the movement of phospholipids from the cytosolic leaflet to the extracellular leaflet (Hankins et al., 2015). ABC transporters play more specialized roles than P4-ATPases, such as transporting molecules across the bilayer alongside the phospholipid (Hankins et al., 2015). In the paper written by Perez et al., the flippase PglK was determined to be an ABC transporter that catalyzed the flipping of lipid-linked oligosaccharides (Perez et al., 2015).
Structure of an ABC Transporter

Figure 6: Ribbon representation of MsbA structure.
Before the discovery of the PgIK flippase in C. jejuni, the MsbA protein found in gram negative E. coli was one of the earliest discovered flippases in bacteria. Until now, crystal structures have only been reported for the MsbA protein, the bacterial flippase of lipid A-core, but no mechanism could be deduced from the structural data (Perez et al., 2015). In addition to the scarcity of structures, there are few reliable in vitro assays that allow the study of native substrate flipping (Perez et al., 2015). MsbA is a flippase protein that plays a crucial role in maintaining the asymmetry of lipids in the cell membrane. The protein is a member of the ATP-binding cassette (ABC) transporter family and it uses energy from ATP hydrolysis to flip lipids from the inner to the outer leaflet of the cell membrane (Eckford and Sharom, 2010).
The main specific lipid that MsbA is known to flip is Lipid A, which is a component of lipopolysaccharide and is mainly responsible for the cytotoxicity of gram negative bacteria (Eckford and Sharom, 2010). The protein has two transmembrane domains, each consisting of six transmembrane helices, that span the cell membrane. The protein also has two nucleotide binding domains (NBDs) that bind and hydrolyze ATP (Eckford and Sharom, 2010). These NBDs are highly conserved regions across different strains. The NBDs are located on the cytoplasmic side of the membrane and they interact with the transmembrane domains to form a functional unit that is responsible for the flippase activity. Studies have revealed that MsbA uses a “power-stroke” mechanism to transport lipids, where the binding of ATP to the NBDs causes a conformational change in the transmembrane domains, which in turn drives the flipping of lipids across the membrane. In a study conducted by Sharom et al, the researchers generated a group of mutant MsbA proteins and discovered that cells with mutant copies resulted in the accumulation of phospholipids and lipid A on the cytoplasmic side of the membrane (Eckford and Sharom, 2010). This demonstrates the essential flippase function that MsbA proteins possess. Such data from MsbA gave researchers insight into the potential structure and mechanism of PglK. However, further research was required to obtain more insight into the protein.
Protein Glycosylation
What is Glycosylation and Why is it Important?
Glycosylation refers to the reaction in which a carbohydrate (i.e. glycan) is attached to another macromolecule, typically a protein (Figure 7) or lipid. It is usually an enzyme-catalyzed reaction, whereas 'glycation' refers to the non-enzymatic addition of sugars to a molecule (Lima et al., 2013). Glycosylation of proteins can occur both during translation and post-translationally, and the attached glycans serve important structural and functional roles in both membrane and secreted proteins (Varki et al., 2009). For instance, glycosylation may be essential to ensuring proper protein folding and stability, as the negatively charged carbohydrate molecules can form stabilizing interactions with positive amino acids (Varki et al., 2009). Similarly, the hydrophilic carbohydrate chain can also increase protein solubility in aqueous environments. Beyond modifying the physicochemical properties of proteins, glycosylation also facilitates important biological functions. Surface glycoproteins serve as attachment points for other cells in a process known as cell-to-cell adhesion. Similarly, bacterial pathogens exploit surface glycoproteins on eukaryotic host cells for attachment, thereby facilitating their colonization and tissue entry (Li et al., 2016). In combat, microbial surface glycoproteins serve as prime targets for the human immune system, as immune cells are able to recognize these sugars utilizing pathogen-recognition receptors (PRRs) (Varki et al., 2009). As pathogens may go on to modify their glycoantigens to evade immune recognition, an understanding of microbial glycosylation is critical in combating such organisms (Li et al., 2016).

Figure 7: Glycoproteins on the cell surface. Glycoproteins are comprised of a sugar component and protein component. They are linked through a glycosidic bond.
Types of Glycosylation
There are three main types of glycosylation:
N-linked glycosylation
N-linked glycosylation involves the addition of a glycan to the amide nitrogen of an asparagine amino acid (Murray & Bender, 2009). In bacteria, N-glycosylation often occurs at the consensus sequence of Asn-Xaa-Ser/Thr (Wacker et al. 2002).
O-linked glycosylation
O-linked glycosylation involves the attachment of glycans to the hydroxyl oxygen of serine, threonine, tyrosine, hydroxylysine, or hydroxyproline side chains (Murray & Bender, 2009).
C-linked glycosylation
C-linked glycosylation is less common and involves the addition of a sugar to a carbon on a tryptophan side chain (Murray & Bender, 2009).
Mechanism of N-Glycosylation
All three domains of life perform N-glycosylation in a similar manner (Figure 8). The sugars are first assembled in the cytoplasm, forming an oligosaccharide precursor (Dell et al., 2010). The sugar is assembled whilst attached via pyrophosphate to a lipid carrier docked in the cell membrane (in bacteria) or endoplasmic reticulum (in eukaryotes). When the sugar is completed, the lipid-linked oligosaccharide aka LLO is flipped from the cytoplasm to the periplasm (in bacteria) or endoplasmic reticulum lumen (in eukaryotes) via the action of a flippase enzyme. Once in their respective cellular location, the oligosaccharide is transferred as a unit from the lipid carrier to its acceptor protein. This step is catalyzed by the oligosaccharyltransferase enzyme (N-OST), found in all domains of life (Dell et al., 2010).
Thus, there are three hallmark events of N-glycosylation (Dell et al., 2010):
- Cytoplasmic assembly of the oligosaccharide on a lipid carrier.
- Flipping of the lipid-linked oligosaccharide across the membrane.
- Transfer of the oligosaccharide to the protein acceptor.
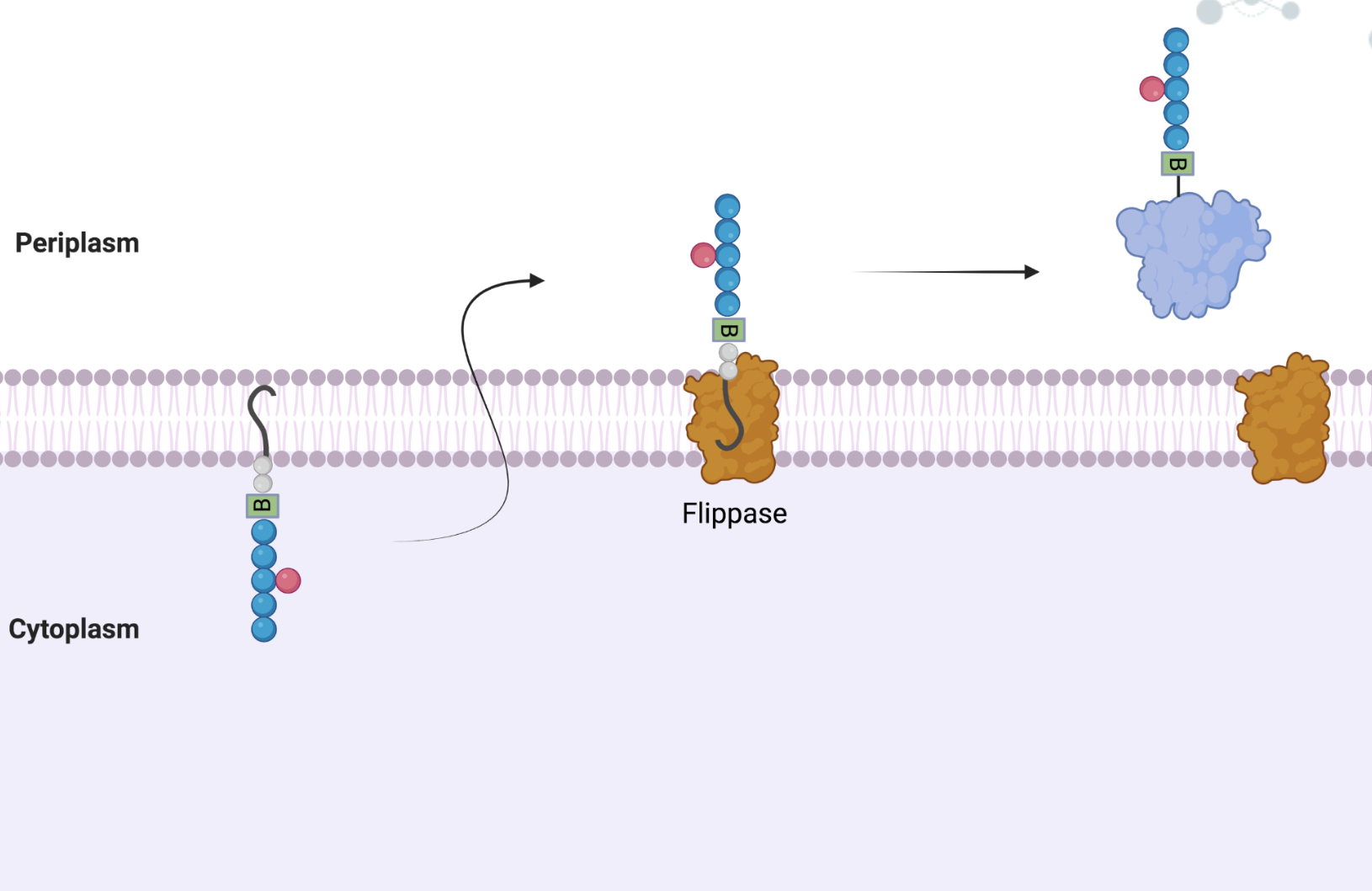
Figure 8: Overview of N-glycosylation. The three key steps of N-glycosylation are shown above. On the left, the lipid linked sugar is assembled. In the center of the image, the flippase translocates the LLO across the membrane, from the cytosolic leaflet to the periplasmic leaflet. On the right, the LLO is transferred to the accepting protein, and thus, the protein is glycosylated.
Lipid-Linked Oligosaccharides
A lipid carrier is a molecule docked in the cell membrane onto which the oligosaccharide is polymerized. Due to its hydrophobicity, it remains in the cell membrane whilst the hydrophilic oligosaccharide component is transferred from the cytoplasm to the periplasm (in bacteria) or endoplasmic reticulum lumen (in eukaryotes). Together, this molecule is known as a lipid-linked oligosaccharide. The lipid carrier component of the LLO differs between the three domains. In eukaryotes and archaea, the lipid is a dolichol. Dolichols are polymers of isoprene units (CH3–C(CH3)=CH–CH2–)n (Dell et al., 2010). In contrast, bacteria use undecaprenyl pyrophosphate as their lipid constituent (Dell et al., 2010). Undecaprenyl is also a polymer of isoprene units; however, it contains 11 isoprenes and possesses one more double bound than the same length dolichol. This double bound is located between carbons 2 and 3 with respect to the alcohol group and decreases the rotational mobility of bacterial LLOs (Dell et al., 2010).

Figure 9: Structure of a lipid linked oligosaccharide. An LLO is composed of polyisoprene, pyrophosphate, and an oligosaccharide. When the three components are polymerized, they form the LLO. The carbon structure of the compound is shown on the right.
The Structure of the Oligosaccharide
Another difference in the N-biosynthetic pathways between eukaryotes and prokaryotes lies in the structure of the oligosaccharide unit. In eukaryotes, the structure of this glycan is highly conserved; in the cytoplasm, the LLO, consists of the heptasaccharide Man5GlcNAc2. After it is flipped from the cytoplasm to the endoplasmic reticulum, the glycan chain is further extended by the addition of 4 mannoses and 3 glucoses, forming Glc3Man9GlcNAc2P-P-Dol (Dell et al., 2010). In contrast, the oligosaccharide is not conserved among bacteria and archaea; each species utilizes varying carbohydrates in their glycosylation processes. For example, Young et al. (2002) identified the structure of the glycan chain present on most proteins in Campylobacter jejuni. Using mass spectrometry and nano-NMR techniques, the structure of the glycan was determined to be heptasaccharide consisting of a linear chain of one D-bacillosamine and five D-N-Acetylgalactosamines with a D-Glucose branch (Young et al., 2002). Bacillosamine is a diaminotrideoxyhexose which is connected to the asparagine residue of proteins (Young et al., 2002).
The structure is as follows:
GalNAc-a1,4-GalNAc-a1,4-[GlucB1,3-]GalNAc-a1,4-GalNAc-a,1,3-Bac-B1.

Figure 10: Structure of the oligosaccharide. The oligosaccharide contains five N-acetylgalactosamines (blue) and one glucose (pink) branch. A di-N-acetyl bacillosamine (green) exists at the end of the sugar, and is linked to asparagine residues.
N-Glycosylation in C. jejuni
Discovery of pgl Locus in C. jejuni
Protein glycosylation was originally thought to be a process that only occurred in eukaryotes, but has now been identified in bacteria and archaea. Bacterial N-glycosylation was first indentified in C. jejuni, a human pathogen that causes gastroenteritis (Alaimo et al., 2006). This post translational modification system is biologically important and is known to be associated with pathogenic processes such as cell to cell recognition and binding; however, its main role is not yet fully understood (Cain et al., 2020). Researchers Wacker et al. (2002) found that the glycosylation system is encoded by the pgl (“Protein Glycosylation”) operon. This operon is found to be highly conserved among the order Campylobacteriales, although N-glycan composition, structure, and pgl gene location may differ between species (Nothaft et al., 2012). Researchers observed that the pgl operon encoded a number of genes with sequence similarities to glycosyltransferases and enzymes required for sugar biosynthesis (Wacker et al., 2002). For example, the pglB gene exhibited strong sequence similarity to the eukaryotic Stt3 protein, a component of the oligosaccharyltransferase complex required for the addition of the glycan chain to eukaryotic proteins (Wacker et al., 2002). Furthermore, the role of each additional glycosyltransferase involved in the synthesis of the glycan chain was elucidated in several subsequent studies (Glover et al., 2005; Linton et al., 2005; Weeparana et al., 2005). As additional evidence for the role of the pgl genes in N-glycosylation, it was found that transfer of the pgl gene cluster to bacterial species that did not possess the ability to N-glycosylate (i.e. E. coli) resulted in the gain of ability to N-glycosylate co-expressed acceptor proteins (Wacker et al., 2002).
Identification of N-Glycoproteins in C. jejuni
In 2002, Linton et al. identified Glycoproteins PEB3 and CgpA as the first non-flagellin glycoproteins in C. jejuni. Furthermore, two distinct protein glycosylation loci were identified in C. jejuni. The first loci is a flagellin modification locus which is involved in the glycosylation of flagellin subunit protein specifically. The second loci identified is the pgl gene locus which is involved in general protein glycosylation. Further research was done regarding PEB3 and it was identified that it was attached to a heptasaccharide through N-linkage at Asn70 (Young et al., 2002). Researchers Wacker et al. (2002) also showed that the heptasaccharide to be added to proteins (GalNAc-a1,4-GalNAc-a1,4-[GlucB1,3-]GalNAc-a1,4-GalNAc-a,1,3-Bac-B1) is first synthesized in the cytoplasm on an undecaprenyl pyrophosphate carrier. At the time of its discovery, researchers hypothesized that the protein PglK was the flippase responsible for transporting this lipid-linked oligosaccharide across the plasma membrane (Wacker et al., 2002). However, its function had never been confirmed.
Confirming the Function of PglK
The flippase function of PglK was confirmed in a 2006 study by Alaimo et al. In doing so, researchers inactivated PglK by inserting a kanamycin resistance cassette into the pglK gene. An experiment was then performed in which R12 antiserum was added to a C. jejuni membrane lysate to identify the presence of glycosylated proteins (Alaimo et al., 2006). R12 reacts with and has a high preference for several C. jejuni glycoproteins, creating a characteristic banding pattern during SDS-PAGE. However, the pglK mutant cells produced a banding pattern that differed from those of the wild type, indicating the absence of several glycosylated proteins (Alaimo et al., 2006). Thus, this hypoglycosylation profile confirmed that pglK encodes a protein involved in N-glycosylation.
To determine whether PglK was an ABC transporter, researchers looked for the presence of an essential ATP-binding domain (NBD) and assessed its functional activity (Alaimo et al., 2006). Conserved amino-acid residues required for the ATPase activity of the ATP-binding domain were mutated, and a specific protein (AcrA) was analyzed for glycosylation (Alaimo et al., 2006). Researchers observed that AcrA was unglycosylated, indicating that PglK requires a functional ATP-binding domain to retain its catalytic activity. On the basis of this observation, researchers concluded that PglK is an ABC transporter (Alaimo et al., 2006).
The final study that confirmed the flippase activity of PglK involved expressing the pgl locus in Escherichia coli. This strain of E. coli lacked a functional wzx gene. This gene encoded a flippase involved in the translocation of O-antigen subunits across the periplasmic membrane for the synthesis of lipopolysaccharide (LPS) (Alaimo et al., 2006). In the absence of the functional Wzx flippase, cells expressing PglK were still able to synthesize LPS (Alaimo et al., 2006). This indicated that PglK had fulfilled the role of a flippase, and it exhibited relaxed substrate specificity in terms of the oligosaccharide it translocated (Alaimo et al., 2006). Altogether, these series of experiments by Alaimo et al. (2006) confirmed the flippase activity of PglK.
Mechanism of N-Glycosylation in C. jejuni
The N-glycosylation pathway in C. jejuni is considered the best-studied system in bacteria (Li et al., 2010). The system is encoded by a 17kb locus called the pgl gene cluster. It contains 12 open reading frames which encode the enzymes responsible for glycosylating over 60 different proteins in C. jejuni (Li et al., 2010). As aforementioned, the heptasaccharide (GalNAc - GalNAc - Glc - GalNAc - GalNAc - GalNAc - diNAcBac) is first assembled on a lipid carrier, undecaprenyl pyrophosphate, to form a lipid-linked oligosaccharide (LLO). The assembly of the heptasaccharide onto the lipid carrier requires five glycosyltransferases (Figure 11).
- PglC: transfers diNAcBac to the lipid carrier
- PglA: adds the second GalNAc residue
- PglJ: adds the third GalNAc residue
- PglH: acts as a polymerase to transfer three GalNAc residues
- PglI: adds the branching Glc residue to complete the heptasaccharide
After assembly in the cytoplasm, the LLO is flipped to the periplasm by the flippase enzyme, PglK (Li et al., 2016). The heptasaccharide is then transferred en bloc (as a unit) to acceptor proteins by the oligosaccharyltransferase PglB (Li et al., 2016).

Figure 11: Mechanism of N-glycosylation in Campylobacter jejuni. PglF, PglE and PglD convert UDP-GlcNAc to UDP-diNAcBac, which is the first sugar residue transferred to UndP carrier by PglC to form UndPP-linked diNAcBac. The glycosyltransferases PglA, PglJ and PglH add five GalNAc residues and one Glucose branch to complete the UndPP-linked heptasaccharide. PglK then flips the completed LLO to the periplasm, where PglB transfers the heptasaccharide to the asparagine residue present in the consensus sequence: Asp/Glc-X1-Asn-X2-Ser/Thr (where X1 and X2 can be any amino acid except proline) (Li et al., 2016).
Transferring the Oligosaccharide to Proteins
Upon PglB-catalyzed transfer of the oligosaccharide to proteins, the N-glycosidic bond between the molecules is formed through nucleophilic attack by the amido group of the acceptor protein. The undecaprenyl pyrophosphate acts as a leaving group following a nucleophilic attack. An amide is not a great nucleophile and thus needs to be “activated” in order to perform the nucleophilic attack (Mohanaty et al., 2020). There are two proposed mechanisms (Figure 12) (Mohanaty et al., 2020):
- A general base deprotonates the amide leading to an immediate tautomer, which is a competent nucleophile to attack the C1 carbon.
- Hydrogen bonds on the amide form that twists the nitrogen atom such that the lone pair is decoupled from conjugation and reactive enough for nucleophilic attack.

Figure 12: Mechanism of N-glycosidic bond formation. (a) General base deprotonation mechanism for nuclepohilic attack from to activate the amide group (b) Hydrogen bonds twisting the N-C of the amide group inorder to increase the nucleophilicity
Both these mechanisms are viable but there has not been any corroborating evidence of any catalytic site that can abstract a proton from the high resolution structures of PglB or any other homologs in the other domains of life (Mohanaty et al., 2020). There is however evidence for the second mechanism. Two residues at the catalytic site, D56 and E319, of PglB are positioned well and are able to form hydrogen bonds with the amide group of Asn (Mohanaty et al., 2020). These residues are also found in AglB and Stt3 the Archaea and Eukaryotic homologs respectively. The formation of these hydrogen bonds leads to the rotation of the N-C bond and thus increases its reactivity. The activated amide attacks the C1 carbon of the LLO thus creating the new N-glycosidic bond which displaces the dolichol-PP carrier. Thus, while the mechanism of PglB was generally well-studied, the mechanism of PglK largely evaded scientists.
Elucidating the Structure and Mechanism of PglK
The only plausible way to definitively determine the mechanism of PglK is through solving its structure. Researchers Perez et al. (2015) tackled this challenge in their 2015 paper. In doing so, crystal structures of PglK uncovered structural features that suggested how the protein flips LLOs from the cytoplasm to the periplasm (Verchère & Menon, 2015). The structural data alongside the proposed mechanism will be outlined in our subsequent Wiki page.
The Importance of PglK and N-Glycosylation
1. N-Glycosylation Mediates Pathogenicity
The findings in this article are significant as C. jejuni is a highly pathogenic bacteria that is responsible for various food-borne diseases, including gastroenteritis (Nothaft & Szymanski, 2010). The pathogen is zoonotic in origin, predominantly found in poultry. Recent studies have shown that C. jejuni contains numerous virulence factors including motility, serum resistance, and adhesion, allowing for bacterial infection (Nothaft & Szymanski, 2010). The proteins associated with these traits contribute to virulence by allowing C. jejuni to adhere and invade human gut epithelial cells (Nothaft & Szymanski, 2010). More than 60 C. jejuni proteins undergo the process of N-linked glycosylation, which impacts the overall pathogenesis of C. jejuni (Nothaft & Szymanski, 2010). When glycosylation fails to occur in these proteins, their ability to aid in infection diminishes significantly. Mutations in the C. jejuni N-glycosylation pathway have been shown to result in reduced chicken and mouse colonization, as well as decreased ability to adhere and invade intestinal epithelial cells (Nothaft & Szymanski, 2010). Further, iron transport and DNA uptake by type IV secretion systems are adversely affected due to loss of N-glycosylation in C. jejuni (Alemka et al., 2013). Thus, understanding the mechanism of N-glycosylation is critical to understanding the organism’s pathogenesis.
2. Immune Evasion
C. jejuni N-glycans also serve as important antigens recognized by immune cells. Research by van Sorge et al. (2009) demonstrated that the heptasaccharide is recognized by human macrophage galactose-type lectin (MGL) (Nothaft & Szymanski, 2010). This dendritic cell receptor captures glycosylated antigens for processing and presentation, initiating an immune signaling cascade that leads to the production of cytokines and recruitment of other immune cells (Nothaft & Szymanski, 2010). C. jejuni pglA mutants which are unable to bind MGL exhibit increased dendritic cell production of the cytokine interleukin-6 (IL-6) (Nothaft & Szymanski, 2010). Thus, it is believed that N-glycosylation of proteins can limit cellular cytokine production to evade host immunity (Nothaft & Szymanski, 2010).
3. Targeting PglK Has Therapeutic Potential
In general, C. jejuni infections are self-limited and acute, however, immunocompromised individuals may face severe symptoms. Illnesses linked with C. jejuni are characterized by diarrhea, abdominal pain, spasms, fever, vomiting, and blood in feces (Stef et al., 2013). Therefore, there is a dire need to combat these infections. Studying the compounds involved in its virulence may be the key to developing a treatment for C. jejuni related diseases. Since PglK is a crucial component of the pathway used to glycosylate these virulence factors (Alemka et al., 2013), loss of the Pglk would impact the overall virulence of the bacteria. Therefore, PglK can be studied in order to develop antivirulence compounds that combat adhesion and invasion into human gut cells. Thus, the elucidation of the PglK structure and mechanism found in this hold grand significance, as they will contribute to future studies that focus on developing these antivirulence compounds.
References
Alaimo, C., Catrein, I., Morf, L., Marolda, C. L., Callewaert, N., Valvano, M. A., Feldman, M. F., & Aebi, M. (2006). Two distinct but interchangeable mechanisms for flipping of lipid-linked oligosaccharides. The EMBO journal, 25(5), 967–976. https://doi.org/10.1038/sj.emboj.7601024
Alemka, A., Nothaft, H., Zheng, J., & Szymanski, C. M. (2013). N-glycosylation of Campylobacter jejuni surface proteins promotes bacterial fitness. Infection and immunity, 81(5), 1674–1682. https://doi.org/10.1128/IAI.01370-12
Cain, J. A., Dale, A. L., Sumer-Bayraktar, Z., Solis, N., & Cordwell, S. J. (2020). Identifying the targets and functions of N-linked protein glycosylation in Campylobacter jejuni. Molecular Omics, 16(4), 287–304. https://doi.org/10.1039/d0mo00032a
Clarke, R. J., Hossain, K. R., & Cao, K. (2020). Physiological roles of transverse lipid asymmetry of animal membranes. Biochimica Et Biophysica Acta (BBA) - Biomembranes, 1862(10), 183382. https://doi.org/10.1016/j.bbamem.2020.183382
Dell, A., Galadari, A., Sastre, F., & Hitchen, P. (2010). Similarities and differences in the glycosylation mechanisms in prokaryotes and eukaryotes. International Journal of Microbiology, 2010, 1–14. https://doi.org/10.1155/2010/148178
Devaux, P. F., Herrmann, A., Ohlwein, N., & Kozlov, M. M. (2008). How lipid flippases can modulate membrane structure. Biochimica et biophysica acta, 1778(7-8), 1591–1600. https://doi.org/10.1016/j.bbamem.2008.03.007
Eckford, P. D., & Sharom, F. J. (2010). The reconstituted Escherichia coli MsbA protein displays lipid flippase activity. The Biochemical journal, 429(1), 195–203. https://doi.org/10.1042/BJ20100144
Graham, T. R. (2021, July 20). Tour de flippase. American Society for Biochemistry and Molecular Biology. Retrieved February 1, 2023, from https://www.asbmb.org/asbmb-today/science/072021/tour-de-flippase
Ho, H., Koth, C. M., & Payandeh, J. (2018). E. coli msba in complex with LPS and inhibitor G092. Protein Data Bank. https://doi.org/10.2210/pdb6bpp/pdb
Kreling, V., Falcone, F. H., Kehrenberg, C., & Hensel, A. (2020). Campylobacter sp.: Pathogenicity factors and prevention methods—new molecular targets for innovative antivirulence drugs? Applied Microbiology and Biotechnology, 104(24), 10409–10436. https://doi.org/10.1007/s00253-020-10974-5
Lima, M., & Baynes, J. W. (2013). Glycation. Encyclopedia of Biological Chemistry, 405–411. https://doi.org/10.1016/b978-0-12-378630-2.00120-1
Lopez-Marques, R. L., Theorin, L., Palmgren, M. G., & Pomorski, T. G. (2014). P4-ATPases: lipid flippases in cell membranes. Pflugers Archiv : European journal of physiology, 466(7), 1227–1240. https://doi.org/10.1007/s00424-013-1363-4
Mohanty, S., Chaudhary, B. P., & Zoetewey, D. (2020). Structural insight into the mechanism of N-linked glycosylation by oligosaccharyltransferase. Biomolecules, 10(4), 624. https://doi.org/10.3390/biom10040624
Murray, R. K., & Murray, R. K. (Robert K. (2009). Harper’s illustrated biochemistry. McGraw-Hill Medical. Nothaft, H., & Szymanski, C. M. (2010). Protein glycosylation in bacteria: Sweeter than ever. Nature Reviews Microbiology, 8(11), 765–778. https://doi.org/10.1038/nrmicro2383
Nothaft, H., Scott, N. E., Vinogradov, E., Liu, X., Hu, R., Beadle, B., Fodor, C., Miller, W. G., Li, J., Cordwell, S. J., & Szymanski, C. M. (2012). Diversity in the protein N-glycosylation pathways within the campylobacter genus. Molecular & Cellular Proteomics, 11(11), 1203–1219. https://doi.org/10.1074/mcp.m112.021519
Patenaude, S. I., Seto N. O. L., Borisova S. N., Szpacenko A., Marcus S. L., Palcic M. M. & Evans S. V. (2002). The structural basis for specificity in human ABO(H) blood group biosynthesis. Nature Structural Biology 9, 685-690. Perez, C. (n.d.). Molecular Mechanism of cell wall membrane proteins. Research Group Camilo Perez - Biozentrum. Retrieved February 1, 2023, from https://www.biozentrum.unibas.ch/research/research-groups/research-groups-a-z/overview/unit/research-group-camilo-perez
Perez, C., Gerber, S., Boilevin, J., Bucher, M., Darbre, T., Aebi, M., Reymond, J. L., & Locher, K. P. (2015). Structure and mechanism of an active lipid-linked oligosaccharide flippase. Nature, 524(7566), 433–438. https://doi.org/10.1038/nature14953
Prof. dr. Kaspar Locher. ETH Zurich. (n.d.). Retrieved February 1, 2023, from https://biol.ethz.ch/en/the-department/people/person-detail.NDkyMzc=.TGlzdC80NjAsOTIzMDMxMjIy.html
Raetz, C. R. H., Reynolds, C. M., Trent, M. S., & Bishop, R. E. (2007). Lipid a modification systems in gram-negative bacteria. Annual Review of Biochemistry, 76(1), 295–329. https://doi.org/10.1146/annurev.biochem.76.010307.145803
Sham, L.-T., Butler, E. K., Lebar, M. D., Kahne, D., Bernhardt, T. G., & Ruiz, N. (2014). Murj is the flippase of lipid-linked precursors for peptidoglycan biogenesis. Science, 345(6193), 220–222. https://doi.org/10.1126/science.1254522 Sharom F. J. (2011). Flipping and flopping–lipids on the move. IUBMB life, 63(9), 736–746. https://doi.org/10.1002/iub.515
Stef, L., Cean, A., Vasile, A., Julean, C., Drinceanu, D., & Corcionivoschi, N. (2013). Virulence characteristics of five new Campylobacter jejuni chicken isolates. Gut pathogens, 5(1), 41. https://doi.org/10.1186/1757-4749-5-41
University of Bern. (n.d.). The Chemical Space Project. Reymond Research Group. Retrieved February 1, 2023, from https://gdb.unibe.ch/research/
van Sorge, N. M., Bleumink, N. M., van Vliet, S. J., Saeland, E., van der Pol, W. L., van Kooyk, Y., & van Putten, J. P. (2009). N-glycosylated proteins and distinct lipooligosaccharide glycoforms of Campylobacter jejuni target the human C-type lectin receptor MGL. Cellular microbiology, 11(12), 1768–1781. https://doi.org/10.1111/j.1462-5822.2009.01370.x
Varki, A., Cummings, R. D., Esko, J. D., Freeze, H. H., Stanley, P., Bertozzi, C. R., Hart, G. W., & Etzler, M. E. (Eds.). (2009). Essentials of Glycobiology. (2nd ed.). Cold Spring Harbor Laboratory Press.
Wacker, M., Linton, D., Hitchen, P. G., Nita-Lazar, M., Haslam, S. M., North, S. J., Panico, M., Morris, H. R., Dell, A., Wren, B. W., & Aebi, M. (2002). N-linked glycosylation in Campylobacter jejuni and its functional transfer into E. coli. Science (New York, N.Y.), 298(5599), 1790–1793. https://doi.org/10.1126/science.298.5599.1790
Why are membrane lipids asymmetrical? MBInfo. (2018, February 6). Retrieved February 1, 2023, from https://www.mechanobio.info/what-is-the-plasma-membrane/why-are-membrane-lipids-asymmetrical/