Table of Contents
A cohort autopsy study defines COVID-19 systemic pathogenesis
Introduction
SARS-CoV-2 is a disease-causing pathogen from the coronaviridae family. It is closely related to SARS-CoV and MERS-CoV, however has lower mortality rates than the two and increased transmission rates. SARS-CoV-2 can lead to severe illness in certain individuals, usually ones who have pre-existing or underlying health conditions. This severe illness is almost always accompanied by a dysfunction in the lungs or other organs (Yao et. al. 2021).
Previously, our group discussed the pathogenesis and transmission of SARS-CoV-2. SARS-CoV-2 causes infection of cell targets by binding a cognate receptor (most commonly angiotensin-converting enzyme 2, or ACE2) with the spike protein and infiltrating the cell cytoplasm, after which it utilizes host cell machinery to create viral proteins for further replication and propagation of viral particles. One key area of inquiry was systemic SARS-CoV-2 infection and the means through which the virus achieves infection of extrapulmonary tissue. Past findings indicate that SARS-CoV-2 passes into the bloodstream 14 days after exposure. From here, the virus may gain access to a variety of body regions, signalling the beginning of systemic SARS-CoV-2 infection presenting a more diverse set of symptoms than in pulmonary infections. Systemic infections present with respiratory symptoms typical of SARS-CoV-2, such as Acute Respiratory Distress Syndrome (ARDS), alongside a range of more diverse symptoms such as immune dysfunction, proteinuria, arrhythmia, diarrhea, impaired consciousness, and sensory deprivation. The threat of multi-organ failure represents severe systemic sequelae to infection, making a case for the necessity of a more robust understanding of how systemic infection is achieved. In this 2021 paper by Yao et al., the range of systemic infection targets are investigated, along with the possible cell targets that may carry the virus to these sites. These findings may inspire future investigations into systemic SARS-CoV-2 infection and possible pharmaceutical interventions.
Rationale
Previous research has shown that older adults with underlying health conditions face an increased risk of severe infection and increased mortality rates due to COVID-19. In this paper, the researchers aimed to discover more about the pathogenesis of SARS-CoV-2 and the changes it can cause in the lungs and the extrapulmonary organs. In this study, the researchers conducted post-mortem autopsies on 26 deceased COVID-19 patients to gain insight into disease progression and systemic infection (Yao et. al. 2021). Since the presence of spike or nucleocapsid protein is considered an indication of SARS-CoV-2 infection, immunohistochemical (IHC) staining of these two proteins is being employed to gauge viral presence in post-mortem tissue. Additionally, single-cell RNA sequencing (scRNA-seq) is being used to detect viral transcripts to determine viral infection of cells (Yao et. al. 2021). By studying post-mortem tissue acquired from these patients, the investigators hoped to better understand SARS-CoV-2 pathology and viral infiltration of extrapulmonary targets.
Study Design
The study was conducted using 26 autopsy cases from four cohorts in Wuhan, China (Yao et al., 2021). The autopsy cases were donated with the consent of the patient’s families from Huoshenshan Hospital, Taikang Tongji Hospital, Zhongfaxincheng Hospital, and Wuhan Jinyintan Hospital (Yao et al., 2021). All deceased individuals died at the start of the pandemic between February 18th and April 4th, 2020. All patients were classified as critically ill COVID-19 patients and the median age was 67.5 years old (Yao et al., 2021). Most patients, around 76.9%, had underlying illnesses, more specifically, lung diseases, chronic cardiovascular diseases, and hypertension (Yao et al., 2021). The major cause of death among the patients was due to severe pulmonary injuries leading to pulmonary failure, pulmonary thromboembolism, dissecting aneurysm rupture, and cardiovascular disorders (Yao et al., 2021). To compare pulmonary effects of COVID-19, control samples of lung tissue were harvested from deceased lung carcinoma or heart failure patients (Yao et al., 2021).
The purpose of the study is to understand SARS-CoV-2 pathology and viral infiltration via various ports of entry of the virus. In order to do so, obtaining tissue samples from pulmonary and extrapulmonary organs where SARS-CoV-2 spread is predicted is vital to study the tissue samples and perform various techniques to confirm the presence of SARS-CoV-2 spike or nucleocapsid proteins among the tissue harvested. Thus, preservation of the tissue is crucial for further study. The method of preservation used was fresh or formalin-fixed, paraffin-embedded, also known as FFPE (Yao et al., 2021). The sample tissue of interest was taken from the autopsy patients and a control tissue for comparison purposes. The tissues are then subject to preservation in formaldehyde for about 18-24 hours (“What Is FFPE Tissue and What Are Its Uses,” 2017). This aims to preserve the tissue structure as well as the proteins present in the tissue itself, which is important when it comes to viral protein detection for SARS-CoV-2 (“What Is FFPE Tissue and What Are Its Uses,” 2017). In formaldehyde, the tissue is dried and washed, and then fixed on a paraffin wax block (“What Is FFPE Tissue And What Are Its Uses,” 2017). Fixation on the block is important since it allows for precise slicing of tissue for examination (“What Is FFPE Tissue and What Are Its Uses,” 2017). FFPE, compared to other preservation methods, was ideal for this study since it allows for the preservation of viral proteins in the sample directly. This is important when using techniques such as immunohistochemistry to identify SARS-CoV-2 spike and nucleocapsid proteins (“What Is FFPE Tissue and What Are Its Uses,” 2017). Other preservation techniques, such as frozen tissue, are more fragile and require special handling procedures compared to FFPE preserved tissue (“What Is FFPE Tissue and What Are Its Uses,” 2017). FFPE is also more cost-effective in the long run and allows for storage of the preserved tissue to be used years later if necessary (Gao et al., 2020). Although FFPE is the preferred method for tissue preservation in this specific study, it has its drawbacks as well. Formaldehyde can form crosslinks with proteins via specific reactions to amino acid groups (O’Rourke and Padula, 2018). This can cause the antigens to not be detected, which can reduce the efficacy and accuracy of antibody binding to antigens during immunohistochemistry (“What Is FFPE Tissue and What Are Its Uses,” 2017). However, certain techniques have been developed over the years to specifically bind to antigens that have crosslinked with formaldehyde (“What Is FFPE Tissue and What Are Its Uses,” 2017). This technique is known as antigen retrieval (“What Is FFPE Tissue and What Are Its Uses,” 2017).
After tissue preservation using FFPE, detection of viral proteins of SARS-CoV-2, which are the spike and nucleocapsid proteins, tissue staining using hematoxylin and eosin is done to visualize the contents of the nucleus and cytoplasm, respectively (Yao et al., 2021). This method is the preferred method and gold standard for tissue staining (Sampias and Rolls, 2022). This method of staining is usually used for FFPE prepared tissues, explaining why it is used in this study specifically. H&E staining is done to view the morphological changes that occur in response to SARS-CoV-2 disease (Sampias & Rolls, n.d.). However, H&E staining is incompatible with immunofluorescence (Fischer et al., 2008). Immunohistochemistry was then performed on the tissue samples using the strep-tavidin-biotin-peroxidase technique with diaminobenzidine (Yao et al., 2021), which is one of the methods used to amplify the target antigen signal to improve sensitivity (“Avidin-Biotin Complex Method for IHC Detection,” n.d.). Immunohistochemistry utilizes antibodies to bind SARS-CoV-2 antigens, which are the spike and nucleocapsid proteins specifically. IHC is used for disease diagnosis to identify cell-specific components, such as proteins or specific biomarkers to identify, diagnose, and categorize disease (“Overview of Immunohistochemistry,” n.d.). Immunohistochemistry allows for the researchers to be able to visualize the port of entry of the virus and the virus being present in physiological barriers, which is what the study is trying to accomplish. The presence of viral proteins was also confirmed via immunofluorescence staining (Yao et al., 2021). This technique is slightly different from IHC in that IF is mainly used to stain a single cell population, such as pulmonary endothelial cells, whereas IHC is used to visualize viral particles in the presence of tissue structure (“IHC or IF: Which is Best for My Study?” 2018). Both techniques are used to identify SARS-CoV-2 in tissues, however, the application of such techniques are slightly different. The differences in both can be seen in Figures 3A and 3B. To confirm that indeed the SARS-CoV-2 spike and nucleoprotein were present in the tissue, immunocytochemistry staining was performed using a transfection protocol (Yao et al., 2021). Transmission electron microscopy (TEM) was used to view tissue morphology due to the high resolution it offers at the nanoscale and even atomic level (“Transmission Electron Microscopy,” n.d.). It is often used in pathology to diagnose diseases in tissue (Creech et al., 2017). By viewing the tissue via TEM, the diffuse alveolar damage was analyzed and characterized, and SARS-CoV-2 spike protein was quantified as well (Yao et al., 2021).
The above methods confirmed SARS-CoV-2 viral protein presence in pulmonary and extrapulmonary tissues. However, further confirmation of SARS-CoV-2 infection was needed, which would be done by detecting viral SARS-CoV-2 nucleic acids to completely confirm that infection occurred in the tissues of interest. Real-time reverse transcription PCR was done in order to detect the viral nucleic acids of SARS-CoV-2 (Yao et al., 2021).
One of the main purposes of the study was to determine the pathological features of SARS-CoV-2 in pulmonary tissue. Although this has been visualized, quantification of the differences between infected lung cells and noninfected cells would need to be investigated to study transcriptional differences between infected and noninfected cells. Thus, single-cell RNA sequencing, known as ScRNA-seq was performed on autopsy case 17 (Yao et al., 2021). Unfixed lung tissues were isolated into single-cell populations to obtain single-cell suspensions (Yao et al., 2021). ScRNA-seq libraries were constructed, pooled, and sequenced using Illumina with 150 base pair paired-end reads (Yao et al., 2021). The BCL files output was converted to FASTQ format, and the data was processed (Yao et al., 2021). After alignment of the reads to a custom genome to detect viral RNA, the results were obtained, normalized, and identified (Yao et al., 2021).
During normalization, clustering, and gene differential expression analysis, the highest quality cells were chosen and normalized (Yao et al., 2021). Now, the cells need to be clustered to compare any differences between them. Clustering is the grouping of similar data points in a data set (Google Developers, 2020). This allows the data points, in this case, the cells, to be grouped together. For this particular study, the researchers selected 2000 variable genes for each cell and formed a KNN graph based on the Euclidean distance (Yao et al., 2021). This is basically a clustering technique in which cells are compared against each other to see which are the closest in distance (Badole, 2021). The distance used here is Euclidean distance, which is the distance between two consecutive points (Badole, 2021). The cells were then clustered and visualized. A Wilcoxon rank-sum test was then done on each cluster to look for any genes that are expressed differently than predicted (Yao et al., 2021). The cell type of each cluster was then marked using the known marker genes (Yao et al., 2021). Clustering of the cells was done in order to find any differentially expressed genes which may function differently after SARS-CoV-2 expression, which is what the study aims to find (Yao et al., 2021). Thus, the whole purpose of the normalization and clustering techniques were to use quantified viral RNA from sequencing techniques and group single cells based on how close they are to each other, to find if there are any genes expressed within the single-cell clusters that are being expressed differently than expected (Yao et al., 2021). This allows for the researchers to be able to see any pathological changes associated with SARS-CoV-2 infection.
Results
Figure 1 - SARS-CoV-2 Organotropism in Patients
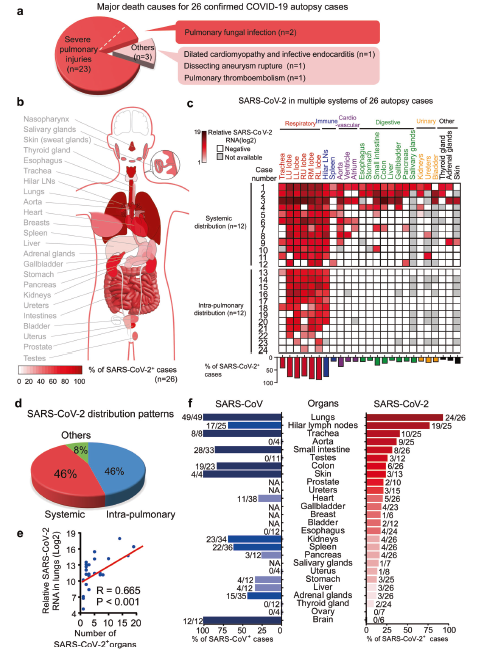
Figure 1. Overview of SARS-CoV-2 Systemic Pathology in COVID-19 Patients. 1a. Causes of death for COVID-19 autopsy patients; 1b. Model of SARS-CoV-2 organotropism; 1c. Heatmap depicting SARS-CoV-2 distribution groups and RNA load in 26 autopsy cases; 1d. Pie chart depicting the percentage of cases in three different SARS-CoV-2 infection groups; 1e. Correlation between SARS-CoV-2 RNA in lungs and number of infected organs; 1f. Comparison of viral infection rates between SARS-CoV-2 based on the current study and SARS-CoV in prior research in postmortem organs from COVID-19 and SARS patients. (Yao et al., 2021)
The paper begins by displaying the results of profiling SARS-CoV-2 organotropism in COVID-19 patients. Out of the 26 autopsy cases examined, the major causes of death included 88% pulmonary fungal infection and 12% other causes, including systemic COVID-19 infection. The other causes included pulmonary thromboembolism, dissecting aneurysm rupture, dilated cardiomyopathy, and infective endocarditis. The researchers also examined different tropisms of SARS-CoV-2 in various tissues in the body. The lungs, heart, kidneys, and lymph nodes displayed the highest percentage of cases where SARS-CoV-2 showed specific tropism in these tissues. The researchers then examined the distribution of SARS-CoV-2 viral RNA in different tissues. The results displayed a higher distribution of viral RNA in the respiratory tissues, however, showed significant evidence for viral RNA presence in extrapulmonary tissues. Several cases showed severe systemic distribution of viral RNA in the pulmonary, immune, cardiovascular, digestive, urinary, and other tissues. The cases of systemic distribution and intrapulmonary distribution were present in equal amounts. The researchers also observed a positive correlation between viral load in the lungs and the presence of SARS-CoV-2 in extrapulmonary organs. This indicates that increasing viral load in the lungs makes individuals more susceptible to systemic infection. The researchers also compared the tropism of SARS-CoV-2 with SARS-CoV. While SARS-CoV showed a high percentage of cases with tropism in the lungs, brain, small intestine, and trachea, SARS-CoV-2 showed a greater tropism for extrapulmonary tissues overall meaning that SARS-CoV-2 is more likely to result in systemic infection (Yao et al., 2021).
Figure 2 - SARS-CoV-2 associated pulmonary pathological changes
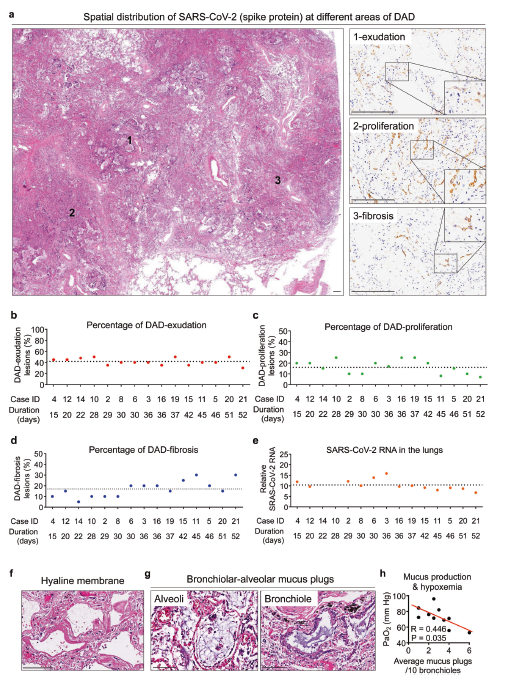
Figure 2. Pulmonary Pathologies in COVID-19 Patients. 2a. H&E and IHC staining showing SARS-CoV-2 Spike protein in pulmonary areas with different features (1, exudation; 2, proliferation; 3, fibrosis) of diffuse alveolar damage (DAD); 2b-e. Proportion of DAD-exudation areas (b), DAD-proliferation areas (c), and DAD-fibrosis areas (d), and the average SARS-CoV-2 RNA in postmortem lungs (e) from 15 COVID-19 autopsy cases; 4f,g. H&E staining reveals hyaline membrane formation (f) and bronchiolar-alveolar mucus (g); 4h. Correlation between average bronchiolar-alveolar mucus plugs and PaO2 level in patients experiencing respiratory disfunction. (Yao et al., 2021)
The paper outlines findings regarding SARS-CoV-2 associated pulmonary pathological changes. Looking at Figure 2A, the spatial distribution of SARS-CoV-2 in different areas of the diffuse alveolar damage is present in pulmonary tissue. The spike protein is observed to exude or leak out from the blood vessels into surrounding pulmonary tissue. The virus is then observed to proliferate in tissue, and then causes fibrosis of pulmonary tissue, meaning scarring and damage of lung tissue. Using the findings regarding pulmonary pathological changes, the researchers create 4 graphs which show the proportion of DAD-exudation areas, DAD-proliferation areas, DAD-fibrosis areas, and average SARS-CoV-2 RNA found in the lungs of 15 out of the 26 autopsy cases in the study, which can be seen in Figures 2B-2E. They also observed the formation of a hyaline membrane formed during diffuse alveolar damage using H&E staining, seen in Figure 2F. H&E staining also showed the presence of mucus plugs in the alveoli and bronchioles of the lungs, which led the researchers to observe the correlation between mucus production and the amount of oxygen present in the lungs, seen in Figures 2G and 2H. The higher the amount of mucus plugs, the lower the oxygen levels in the alveoli, leading to a negative correlation (Yao et al., 2021).
Figure 3 - The presence of SARS-CoV-2 in the endothelium of physiological barriers in the lungs, kidneys, and testes
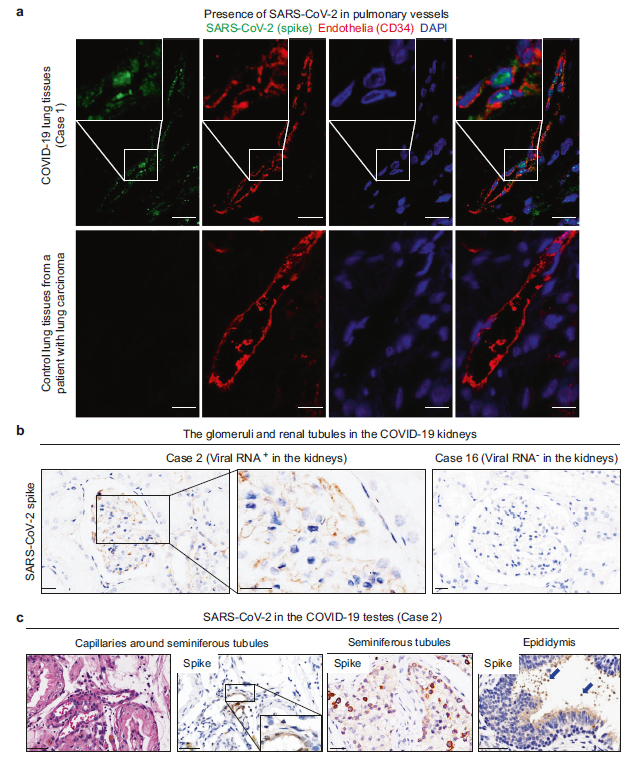
Figure 3. Infection of Endothelia by SARS-CoV-2. 3a. Immunofluorescent staining of SARS-CoV-2 spike protein and CD34 in endothelia of COVID-19 lung tissues and control lung tissues from a patient with lung carcinoma; 3b. IHC staining reveals Spike protein in glomeruli with filtrated endothelium barriers in post-mortem kidneys positive for SARS-CoV-2. Uninfected kidney used for control; 3c. H&E staining and IHC staining showing SARS-CoV-2 spike in endothelia of blood-testis barrier, seminiferous tubules, and sperms in the epididymis of testes from COVID-19 autopsy patients (Yao et al., 2021)
One major idea the paper explored was finding the presence of SARS-CoV-2 in physiological barriers of pulmonary and extrapulmonary tissues. The researchers wanted to see if SARS-CoV-2 spike and nucleocapsid proteins were present in a certain tissue and if they were able to cross any physiological barriers. They began by looking for evidence of SARS-CoV-2 in the endothelium of the lungs by examining two cases, a patient with COVID-19, and a patient who has tested negative for COVID-19 but is afflicted with lung carcinoma (Figure 3A). In Case 1, the presence of SARS-CoV-2 in the lungs is detected using immunofluorescent staining. Immunofluorescent staining was used to see where CD34, a transmembrane phosphoglycoprotein that acts as facilitator of cell migration, is present on pulmonary endothelial cells (AbuSamra et al., 2017). DAPI was used to stain the cell nuclei to assess cell morphology (Tarnowski et al., 1991). Comparing the images of Case 1 to Case 2 in Figure 3A, it can be observed that the pulmonary vessels in Case 1 are severely constricted, especially in areas where SARS-CoV-2 is highly concentrated. In contrast, the pulmonary vessels of control patients with lung carcinoma in Case 2 are not constricted like those of COVID-19 patients. This can be an interesting observation that can be further studied to better understand this information.
As evidence for SARS-CoV-2 presence in the lungs emerged, the researchers also wanted to find evidence of SARS-CoV-2 systemic dissemination in extrapulmonary tissues as well, such as the kidneys and testes. Through their experimentation, they found evidence of viral proteins in parts of the kidneys. By examining Figure 3B, the presence of SARS-CoV-2 in the glomerulus and renal tubules of the kidneys using immunohistochemical staining is evident. It was mainly localised in the filtration barriers, as well as in epithelial cells of the proximal convoluted tubule. An interesting next step in this scenario would be to study the potential impacts of SARS-CoV-2 infection on the kidneys to see if it has any effect on vital functions, such as the glomerular filtration rate.
A closer look at SARS-CoV-2 systemic dissemination across physiological barriers revealed an unusual discovery: SARS-CoV-2 viral proteins in the testes. Although it was expected to see these viral proteins in pulmonary tissue and some extrapulmonary tissue such as the kidneys, the testes proved to be a surprising target for SARS-CoV-2 organotropism. Looking at Figure 3C, the presence of SARS-CoV-2 in the endothelium of the blood-testis barrier, seminiferous tubules, and sperms in the epididymis of the testes can be seen using immunohistochemical staining and H&E staining (Yao et al., 2021). The images suggest that SARS-CoV-2 is able to penetrate physiological barriers leading to organotropism of the virus to specific physiological barriers of the body.
Figure 4: SARS-CoV-2 infects circulating and infiltrating macrophages and monocytes
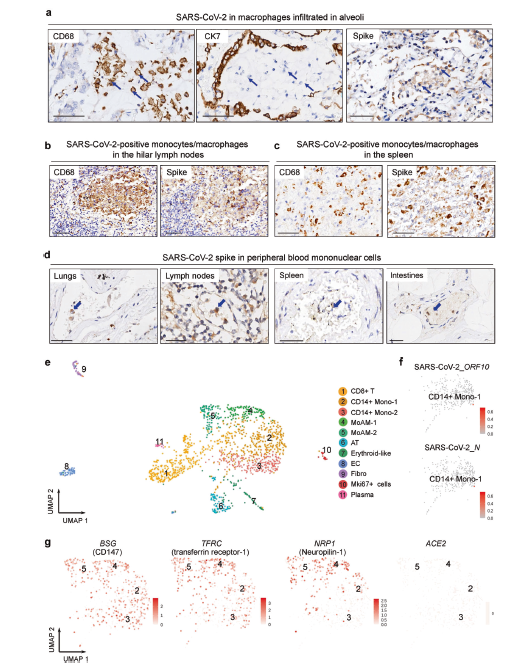
Figure 4a. Evidence of SARS-CoV-2 Infection in Immune Cells. 4a. IHC staining reveals CD68, CK7 and Spike protein in alveolar exudates in blue. Arrows point towards macrophages; 4b. & 4c IHC staining reveals CD68 and Spike protein on macrophages/monocytes in serial sections of spleen and lymph nodes; 4d. IHC staining reveals Spike protein on peripheral blood mononuclear cells in blue. Serial sections obtained from lungs, lymph nodes, spleen, and intestines; 4e. UMAP plot displays scRNA-seq results of 1437 cells from autopsy lung tissue; 4f. Detection of SARS-CoV-2 transcripts for ORF 10 or Nucleocapsid genes; UMAP showing detection of CD147, transferrin-receptor 1, neuropilin-1, and ACE2 transcripts via scRNA-seq (Yao et al., 2021)
Circulating macrophages and monocytes represent a very promising means of systemic pathogenesis. In order to confirm SARS-CoV-2 infection of alveolar macrophages, alveolar exudate was imaged using IHC staining for spike protein, CK7, and CD68. The resulting images revealed predominantly the presence of CD68 macrophages bearing the SARS-CoV-2 spike protein (Fig. 4a) (Yao et al., 2021).
IHC staining of serial sections revealed CD68 macrophages and monocytes bearing SARS-CoV-2 spike protein in the lymph nodes (Fig. 4b) and spleen (Fig. 4c). IHC stained serial sections also revealed SARS-CoV-2 spike protein in peripheral mononuclear cells in lungs, kidneys, intestines, lymph nodes, and spleen (Fig. 4d). The presence of macrophages/monocytes bearing viral spike protein in alveolar and extrapulmonary tissue strongly suggests infiltrating immune cells as a means of viremic, followed by systemic, infection (Yao et al., 2021).
Single-cell RNA sequencing of lung tissue from autopsy case 17 isolated 2 hours after death revealed the presence of CD14+ macrophages and monocyte-derived alveolar macrophages (MoAMs) (Fig. 4e). The presence of SARS-CoV-2 open-reading frame (ORF) 10 transcripts in CD14+ monocytes confirms SARS-CoV-2 infection of pulmonary monocytes (Fig. 4f) (Yao et al., 2021).
Macrophages and monocytes have demonstrated themselves to be important SARS-CoV-2 infection targets. However, macrophages have low expression of the SARS-CoV-2 spike protein cognate receptor, angiotensin-converting enzyme 2 (ACE2) (Ren et al., 2021). To understand if other identified SARS-CoV-2 receptors mediated viral entry into macrophages and monocytes, mRNA of BSG (encoding CD147), TFRC (encoding transferrin receptor-1), and NRP1 (encoding neuropilin-1) in CD14+ monocytes/macrophages and MoAMs were detected using scRNA-seq and plotted via UMAP. CD14+ monocytes and MoAMs expressed all three of the aforementioned: BSG, TFRC, and NRP1. However, ACE2 was not expressed by these cells, implying that viral entry into macrophages/monocytes may be mediated by CD147, transferrin receptor-1, or neuropilin-1 (Fig. 4g) (Yao et al., 2021).
The sum of these findings indicates that SARS-CoV-2 achieves systemic dissemination to target sites such as the kidneys and intestines through infection of circulating and infiltrating macrophages and monocytes.
Discussion & Future Steps
Yao et al. have demonstrated the range of susceptible organs in systemic SARS-CoV-2 infections. By doing so, the risks incurred by viremia, and their clinical significance, is more clearly understood. The authors have also proposed a possible vehicle for viremic transmission by showcasing infection of circulating and infiltrating macrophages found in organs implicated in systemic SARS-CoV-2.
A prominent facet of SARS-CoV-2 pathology that remains poorly understood is its mechanisms of systemic infection. Of chief importance is understanding how SARS-CoV-2 makes its way from pulmonary to extrapulmonary tissue. Once the virus arrives in the vicinity of other organs, especially those protected by physiological barriers, it is pivotal to understand how infection is initiated.
Study findings indicate evidence of viral infection in the endothelia of physiological barriers, raising the possibility of invasion of physiological barriers being a prerequisite for systemic infection. Additionally, SARS-CoV-2 infection of vascular endothelium carries the risk of vasculitis, or inflammation of blood vessels, which has the potential to implicate other organs and tissue in downstream effects via blood/oxygen deprivation. Vasculitis in SARS-CoV-2 must bear further investigation, alongside its role in propagating systemic symptoms of SARS-CoV-2 in organs not infected by the virus. Another implication of endothelial infections concerns thrombosis, one of the hallmark features of severe COVID-19, which, in turn, is followed by ischemia. Infection of and damage to vascular endothelium may be a potential cause of such symptoms. These findings indicate significant risk to those with conditions such as diabetes or hypertension, which are marked by endothelial damage (Bonetti, 2003).
Yao et al. confirmed that SARS-CoV-2 infection results in differences in macrophages composition, leading to a reduction of alveolar resident macrophages and an increase in monocyte-derived macrophages with pro-inflammatory phenotypes. These proinflammatory macrophages are found at various distant infection sites. Some of the multiorgan effects and diversity of symptoms may be explainable by the activity of proinflammatory macrophages. For instance, lymphopenia, a common immune dysfunction observed in COVID-19 patients, may result from macrophages phagocytizing lymphocytes in the lymph nodes (Huang et al., 2020). Another implication of macrophages serving as a vehicle of systemic pathogenesis is the risk of fibrosis. Past studies have determined that a subset of macrophages has demonstrated a capacity for tissue repair and fibrosis in response to damage, which may limit the tissue surface area available for essential bodily functions in infected organs, such as filtration in the kidneys (Wauter et al., 2020). Indeed, a similar phenomenon is observed in respiratory infections via pulmonary fibrosis permanently decreasing surface area for gas exchange.
These findings also hold important implications for antibody-based COVID-19 therapies. One of the possible means through which macrophage infection may occur is by phagocytosis, followed by an escape from the lysosome into the cytoplasm. If this is the case, antibody-based therapies that promote uptake by macrophages may potentially enhance disease progression (Arvin et al., 2020).
Infection of extrapulmonary tissue was confirmed via scRNA-seq detection of SARS-CoV-2 ORF 10 transcripts in CD14+ monocytes and macrophages infiltrating sites such as the kidneys, spleen, intestines, and testes. These findings, combined with the multi-organ failure that characterized most fatalities in the systemic cohort, confirm SARS-CoV-2 systemic infection and pathology. As macrophages represent a critical, and mobile, immune cell population, understanding viral entry mechanisms for macrophage infection is paramount. Macrophages/monocytes rarely express ACE2 and TMPRSS2 (Ren et al., 2021). The study findings reveal alternative SARS-CoV-2 receptors - CD147, transferrin-receptor 1, and neuropilin-1 - on macrophages, the distribution of which correlates with SARS-CoV-2 infection. This finding has value in determining the course for future pharmaceutical research, especially for pharmacological solutions to limiting systemic SARS-CoV-2 infection via binding inhibition. However, it is necessary to confirm that these proteins indeed allow for cell infection in the case of macrophages. This study has not demonstrated that SARS-CoV-2 achieves viral entry by interacting with CD147, transferrin-receptor 1, or neuropilin-1. Future studies will need to demonstrate the interaction between spike and these proteins on macrophages, as well any differences that may exist in their capacity to facilitate infection.
What remains unknown are the precise mechanisms through which SARS-CoV-2 gains access to infection sites protected by physiological barriers. Nucleocapsid and spike protein has been found in barrier endothelia of infected organs, such as the kidneys and testes, implying that endothelial infection is a condition to accessing these sites, but the mechanisms involved remain unclear. Another matter of uncertainty arises from the study’s applicability to younger populations. With an average age of 67.5 among the cohort, it is possible symptoms of a systemic infection may differ in younger patients. In addition, the presence of comorbidities - which may be especially pronounced in an older cohort - and secondary infections acquired during the disease course make discerning the cause of death difficult. Finally, with the above limitations, it is difficult to be certain that the full range of infection targets has been represented by this study. As such, follow-up studies with more diverse, and healthier, cohorts with fewer comorbidities will be instrumental in gaining a clearer understanding of SARS-CoV-2 systemic pathogenesis.
References
AbuSamra, D. B., Aleisa, F. A., Al-Amoodi, A. S., Ahmed, H. M. J., Chin, C. J., Abuelela, A. F., Bergam, P., Sougrat, R., & Merzaban, J. S. (2017). Not just a marker: CD34 on human hematopoietic stem/progenitor cells dominates vascular selectin binding along with CD44. Blood Advances, 1(27), 2799–2816. https://doi.org/10.1182/BLOODADVANCES.2017004317
Arvin, A. M., Fink, K., Schmid, M. A., Cathcart, A., Spreafico, R., Havenar-Daughton, C., Lanzavecchia, A., Corti, D., & Virgin, H. W. (2020). A perspective on potential antibody-dependent enhancement of SARS-COV-2. Nature, 584(7821), 353–363. https://doi.org/10.1038/s41586-020-2538-8
Avidin-Biotin Complex Method for IHC Detection. (n.d.). Thermo Fisher Scientific. Retrieved March 2, 2022, from https://www.thermofisher.com/ca/en/home/life-science/protein-biology/protein-biology-learning-center/protein-biology-resource-library/pierce-protein-methods/avidin-biotin-complex-method-ihc-detection.html#5
Badole, M. (2021, August 6). How KNN Uses Distance Measures? Analytics Vidhya. Retrieved March 2, 2022, from https://www.analyticsvidhya.com/blog/2021/08/how-knn-uses-distance-measures/
Bonetti, P. O., Lerman, L. O., & Lerman, A. (2003). Endothelial dysfunction. Arteriosclerosis, Thrombosis, and Vascular Biology, 23(2), 168–175. https://doi.org/10.1161/01.atv.0000051384.43104.fc
Creech, M. K., Wang, J., Nan, X., & Gibbs, S. L. (2017). Superresolution Imaging of Clinical Formalin Fixed Paraffin Embedded Breast Cancer with Single Molecule Localization Microscopy. Scientific Reports 2017 7:1, 7(1), 1–10. https://doi.org/10.1038/srep40766
Fischer, A. H., Jacobson, K. A., Rose, J., & Zeller, R. (2008). Hematoxylin and eosin staining of tissue and cell sections. CSH Protocols, 2008(5). https://doi.org/10.1101/PDB.PROT4986
Gao, X. H., Li, J., Gong, H. F., Yu, G. Y., Liu, P., Hao, L. Q., Liu, L. J., Bai, C. G., & Zhang, W. (2020). Comparison of Fresh Frozen Tissue With Formalin-Fixed Paraffin-Embedded Tissue for Mutation Analysis Using a Multi-Gene Panel in Patients With Colorectal Cancer. Frontiers in oncology, 10, 310. https://doi.org/10.3389/fonc.2020.00310
Google Developers. (2020). What is Clustering? Google. Retrieved March 2, 2022, from https://developers.google.com/machine-learning/clustering/overview
Huang, C., Wang, Y., Li, X., Ren, L., Zhao, J., Hu, Y., Zhang, L., Fan, G., Xu, J., Gu, X., Cheng, Z., Yu, T., Xia, J., Wei, Y., Wu, W., Xie, X., Yin, W., Li, H., Liu, M., … Cao, B. (2020). Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. The Lancet, 395(10223), 497–506. https://doi.org/10.1016/s0140-6736(20)30183-5
IHC or IF: Which is Best for My Study? (2018, June 28). StageBio. Retrieved March 2, 2022, from https://www.stagebio.com/blog/ihc-or-if-which-is-best-for-my-study
O’Rourke, M. B., & Padula, M. P. (2018). Analysis of formalin-fixed, paraffin-embedded (FFPE) tissue via proteomic techniques and misconceptions of antigen retrieval. BioTechniques, 60(5), 229–238. https://doi.org/10.2144/000114414/ASSET/IMAGES/LARGE/TABLE1.JPEG
{kind=link}
Overview of Immunohistochemistry. (n.d.).Thermo Fisher Scientific. Retrieved March 2, 2022, from https://www.thermofisher.com/ca/en/home/life-science/protein-biology/protein-biology-learning-center/protein-biology-resource-library/pierce-protein-methods/overview-immunohistochemistry.html
Ren, X., Wen, W., Fan, X., Hou, W., Su, B., Cai, P., Li, J., Liu, Y., Tang, F., Zhang, F., Yang, Y., He, J., Ma, W., He, J., Wang, P., Cao, Q., Chen, F., Chen, Y., Cheng, X., … Zhang, Z. (2021). Covid-19 immune features revealed by a large-scale single-cell transcriptome Atlas. Cell, 184(7). https://doi.org/10.1016/j.cell.2021.01.053
Sampias, C., & Rolls, G. (n.d.). An Intro to H&E Staining: Protocol, Best Practices, Steps & More. Leica Biosystems. Retrieved March 2, 2022, from https://www.leicabiosystems.com/en-ca/knowledge-pathway/he-staining-overview-a-guide-to-best-practices/
Tarnowski, B. I., Spinale, F. G., & Nicholson, J. H. (1991). DAPI as a useful stain for nuclear quantitation. Biotechnic & histochemistry : official publication of the Biological Stain Commission, 66(6), 297–302.
Transmission Electron Microscopy. (n.d.). Thermo Fisher Scientific. Retrieved March 2, 2022, from https://www.thermofisher.com/ca/en/home/electron-microscopy/products/transmission-electron-microscopes.html
Wauters, E., Van Mol, P., Garg, A. D., Jansen, S., Van Herck, Y., Vanderbeke, L., Bassez, A., Boeckx, B., Malengier-Devlies, B., Timmerman, A., Van Brussel, T., Van Buyten, T., Schepers, R., Heylen, E., Dauwe, D., Dooms, C., Gunst, J., Hermans, G., Meersseman, P., … Lambrechts, D. (2020). Discriminating mild from critical COVID-19 by innate and adaptive immune single-cell profiling of bronchoalveolar lavages. https://doi.org/10.1101/2020.07.09.196519
What Is FFPE Tissue And What Are Its Uses. (2017, August 30). BioChain Institute Inc. Retrieved March 2, 2022, from https://www.biochain.com/blog/what-is-ffpe-tissue-and-what-are-its-uses/
Yao, X.-H., Luo, T., Shi, Y., He, Z.-C., Tang, R., Zhang, P.-P., Cai, J., Zhou, X.-D., Jiang, D.-P., Fei, X.-C., Huang, X.-Q., Zhao, L., Zhang, H., Wu, H.-B., Ren, Y., Liu, Z.-H., Zhang, H.-R., Chen, C., Fu, W.-J., … Bian, X.-W. (2021). A cohort autopsy study defines COVID-19 systemic pathogenesis. Cell Research, 31(8), 836–846. https://doi.org/10.1038/s41422-021-00523-8