Table of Contents
SARS-CoV-2 Infection Generates Tissue-localized Immunological Memory in Humans
Introduction
Virus-specific circulating memory T cells, B cells, and antibodies
SARS-CoV-2 infection induces an adaptive immune response in the body allowing T and B virus-specific cells to eliminate the virus in the respiratory tract. Protection from re-infection is associated with the generation of memory lymphocytes that continue to circulate the peripheral blood up to a year after the initial infection (Dan et al., 2021). The memory T cells are generated from exposure of naïve T cells to the virus, which then produces a population of effector T cells. The effector T cells either undergo apoptosis following viral clearance, or they can differentiate into antigen-specific memory T cells. The central and effector memory T cells circulate through the blood, as well as entering lymphoid tissues. Whereas, the tissue-resident memory T cells are restricted to the peripheral tissues where re-infection is likely to occur. These subsets of memory T cells patrol the body and wait to be activated upon viral re-entry into the infection sites. Memory T cells further differentiate into CD4+ T cells which can activate B cells, and CD8+ T cells which cytotoxically kill infected cells (Gray et al., 2018). SARS-CoV-2 specific memory T cells were detected in circulation 6 months after infection (Zuo et al., 2021). However, studies have not retained virus-specific cells from the peripheral tissues and organs of patients infected with COVID-19. In coordination with memory T cells, memory B cells are created in the lymphoid organs from naïve B cell interactions with follicular helper T cells germinal centers. The memory B cells are primed for optimal viral detection and response through isotype switching and somatic hypermutation in the germinal center. The relationship between memory B and T cells in the peripheral tissues requires further understanding.
Neutralizing antibodies which act against the viral spike protein of SARS-CoV-2 are another form of protection introduced by vaccines. SARS-Cov-2 infection was shown to generate spike, receptor binding domain and nucleocapsid specific memory B cells in the peripheral blood (Dan et al., 2021). Previous studies have been confined to examining immune responses in the peripheral blood, but Poon et al., extracted memory cells from diverse tissues of donors including the lymphoid organs (Poon et al., 2021)
The memory CD4+ T cells, memory CD8+, memory B cells, and antibodies are the four aspects of immunological memory which contribute to protective immunity against COVID-19. Although patients display heterogenous adaptive immune responses, 90% of patients sustain immune memory to SARS-CoV-2 6 months after initial infection (Dan et al., 2021).
Localized protection in the lungs and lymph nodes
Memory lymphocytes are found in the lymphoid tissues, circulation, and also localize at infection sites/peripheral tissues. Previous mouse model studies have observed tissue resident memory T cells (Trm) mediate optimal protective responses to respiratory infections when residing in the lungs and lung-associated lymph nodes (Teijaro et al., 2011). The non-circulating Trm cells were CD4+ and displayed higher protection to respiratory infections, than the spleen-derived memory T cells (Teijaro et al., 2011). These results demonstrate the possibility of a new class of CD4+ memory T cells retained in the lung tissue compartments, which are an important factor of protection at the mucosal sites (Teijaro et al., 2011). Another study demonstrated effector memory T cells and tissue-resident memory T cells were detectable in the airways of patients with severe COVID-19 (Liao et al., 2020). They also identified severe COVID-19 was associated with high levels of proinflammatory macrophages in the bronchoalveolar lavage fluid, while moderate cases had high levels of clonally expanded CD8+ T cells (Liao et al., 2020). There is evidence that tissue-resident memory B cells in the lungs and lung-associated lymph nodes are important for protection from secondary respiratory infections (Allie et al., 2018). The detected tissue-resident memory B cells were formed early after infection and relied on continuous antigen encounter within the lung tissues (Allie et al., 2018). They were also functionally distinct from circulating B cells in their ability to accelerate the antibody-secreting response upon secondary infection (Allie et al., 2018). Overall, immunological memory following SARS-CoV-2 infection consists of localized protection in the lungs and lung-associated lymph nodes.
Purpose
Poon et al., examined SARS-CoV-2 seropositive organ donors to determine whether memory CD4+ T, CD8+ T, and B cells were generated and sustained in response to infection. They aimed to further understand the breadth of virus-specific T and B cell memory, so improved vaccine strategies could be developed.
Key Findings
- SARS-CoV-2 specific memory CD4+ T cells, memory CD8+ T cells, and memory B cells were predominantly localized in the lungs and lung-associated lymph nodes 6 months following infection (Poon et al., 2021).
- SARS-CoV-2 specific germinal center B cells and follicular helper T cells were also found in the lung-associated lymph nodes which is a result of persistent germinal center reactions following infection (Poon et al., 2021).
- SARS-CoV-2 specific immunological memory is defined by the coordination of cellular and humoral immunity in the tissues (Poon et al., 2021).
Results
Organ donor cohorts for analysis of SARS-CoV-2 specific immune responses
| Donor | Age | Blood | BM | Spleen | Lung | LLN | GLN |
|---|---|---|---|---|---|---|---|
| D492 | 74 | ✅ | ✅ | ✅ | ✅ | ✅ | |
| D495 | 42 | ✅ | ✅ | ✅ | ✅ | ✅ | |
| D498 | 71 | ✅ | ✅ | ✅ | ✅ | ✅ | |
| HDL113 | 10 | ✅ | ✅ | ✅ | ✅ | ✅ |
Table 1: Seropositive donors and their respective tissue contributions. D492, 495, and 498 are all adult donors, while HDL113 represents the pediatric donor. BM = bone marrow; LLN = lung-associated lymph node; GLN = gut-associated lymph node.
The study began by obtaining human tissues from deceased organ donors (Poon et al., 2021). Because the study sought to analyze the natural memory response generated in response to SARS-CoV-2 infection, the researchers required tissues from previously infected, but unvaccinated donors (Poon et al., 2021). These donors also must have succumbed to non-infectious causes (Poon et al., 2021).
To obtain the necessary organ and tissue samples, the group collaborated with an organ procurement organization, or OPO (Poon et al., 2021). OPOs are non-profit organizations responsible for the recovery of organs from deceased donors for transplantation (UNOS, 2022; Poon et al., 2021). In the United States, there are 57 such organizations, and in this study, the tissue and organ samples were procured through the OPO LiveOnNY (UNOS, 2022; Poon et al., 2021).
| T Cell Type | Cell-Surface Markers |
|---|---|
| CD4+ | OX40, 4-1BB, CD40 ligand (CD40L) |
| CD8+ | CD25, 4-1BB |
| TCM (T effector memory) | CD45RA-, CCR7 |
| TEM (T central memory) | CD45RA-, CCR7- |
| TEMRA (terminally differentiated effector memory T) | CD45RA+, CCR7- |
| stem-like | CD45RA, CCR7 |
| TRM (resident memory T cells) | CD69, CD103 |
Table 2: Cell surface receptors used by Poon et al. to isolate specific subpopulations of T cells.
The donors were tested for an ongoing SARS-CoV-2 infection and were confirmed PCR-negative for the virus (Poon et al., 2021). Donor samples either had a documented medical history of previous COVID-19 infection and/or were seropositive against the implicated virus, having IgG against the N, S, and RBD, as well as broadly neutralizing SARS-CoV-2-specific antibodies (Poon et al., 2021).
In total, the experimental cohort comprised four individuals from whom organ and tissue samples were obtained (Poon et al., 2021). Three of the donors were adults, and one was pediatric (Poon et al., 2021). Table 1 outlines the specifics of each individual’s tissue contribution (Poon et al., 2021). Overall, the group obtained samples of: lung-associated lymph node (LLN), gut-associated lymph node (GLN), lung, and spleen tissue, as well as blood and bone marrow samples (Poon et al., 2021). Seven individuals comprised the control group; these samples were collected from pre-pandemic seronegative organ donors who had also succumbed to non-infectious causes (Poon et al., 2021). Tissue samples were processed to obtain live mononuclear cells, including leukocytes, which were then subjected to further testing (Poon et al., 2021).
SARS-CoV-2-specific T cells in lung and lymphoid tissues
To identify T cells specific to SARS-CoV-2, the group conducted an activation-induced marker (AIM) assay, whereby they co-incubated the isolated leukocytes from different tissues with peptide megapools (MPs) (Poon et al., 2021; Reiss et al., 2017). These MPs are large collections of peptides derived from the fragmentation of the S protein, as well as peptides synthesized based on predicted CD4+ and CD8+ T cell epitopes for HLA class I and II (Poon et al., 2021). If a T cell binds these SARS-CoV-2-specific peptides, then it must express the cognate receptor, called an activation induced marker, or AIM (Poon et al., 2021).
Following this procedure, the cells were then subjected to flow cytometry (Poon et al., 2021). The combination of AIMs used to identify the different classes of T cells are shown in Table 2 (Poon et al., 2021). From there, they then sought to understand which cells were expressing which types of T cell predominately (Poon et al., 2021). Figure 1 illustrates their observations for the tissue-specific expression of CD4+ and CD8+ T cells (Poon et al., 2021). The majority of these T cells were specific to the S protein epitopes rather than to the class I and II HLA epitopes (Poon et al., 2021).

Figure 1: Tissue expression of SARS-CoV-2-specific T cells (Poon et al., 2021). Figure 1A shows the overall SARS-CoV-2 specific CD4+ T cell responses (Poon et al., 2021). In general, there was significantly greater expression of CD4+ T cells in all tissues studied except the spleen (Poon et al., 2021). There was especially high expression of the T cells in the bone marrow (BM) and gut- and lymph-associated lymph nodes (GLN and LLN, respectively) (Poon et al., 2021). As illustrated in Figure 1B, there was generally a lower magnitude expression of SARS-CoV-2-specific CD8+ T cells (Poon et al., 2021). The experimental cohort is represented in red, while the seronegative control group is represented in blue. Overall, lung-associated lymph nodes were the dominant site of virus-specific CD8+ and CD4+ T cells (Poon et al., 2021). The circle represents donor D492, the square represents D495, the upward-pointing triangle represents D498, and the inverted triangle represents pediatric donor HDL113.
SARS-CoV-2-specific T cells are maintained in tissues as circulating and resident memory subsets
Next, the group sought to understand the expression of SARS-CoV-2-specific memory T cells in the different tissues (Poon et al., 2021). Figure 2 summarizes the results observed for CD4+ and CD8+ T cells (Poon et al., 2021). For the CD4+ T cells, the majority were TEM and TEMRA cells, the former of which dominates in all but the lymph nodes (Poon et al., 2021).
To find tissue-resident memory T cells (TRM), flow cytometry was used to isolate CD4+ and CD8+ T cells that co-express residency markers CD69 and CD103 (Poon et al., 2021). They found that most of these cells were found in the lung, while some CD8+ TRMs were also found in the lymph nodes (Poon et al., 2021). In summary, the group found that memory T cells are distributed throughout the body, and most TRMs are found in the lungs (Poon et al., 2021).
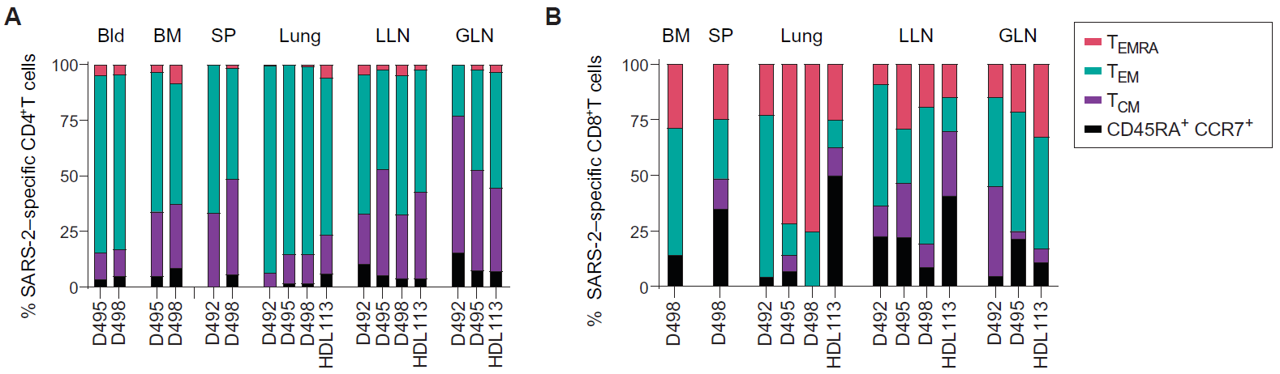
Figure 2: Tissue expression of SARS-CoV-2-specific CD4+ and CD8+ memory T cells among donors. Figure 2A shows the relative contribution of each T cell type to a given tissue profile. In general, T effector memory cells are predominant, followed in contribution by T central memory cells, terminally differentiated effector (EMRA) T cells, and stem-like CD45RA+CCR7+ cells. Figure 2B shows the relative contribution of CD8+ T cells by tissue. Most CD8+ T cells observed were effector memory and EMRA. Notably, the pediatric donor harbored a greater proportion of stem-like T cells in the lungs and LLNs. Figure from Poon et al., 2021.
Tissue specificity and heterogeneity of functional responses to SARS-CoV-2
Heterogeneity exists in functional responses among individuals and sites assessed by quantification of 50 immune mediators produced in different tissue sites (Poon et al., 2021). As seen in Figure 3, for donor D492, the specific functional responses to SARS-CoV-2 were primarily distributed in the lung (Poon et al., 2021). For donor D495, the same responses occur in the lung lymph nodes (LLN) and blood (Poon et al., 2021). For donor D498 and HDL113, various functional responses among different sites were observed (Poon et al., 2021). Donors D492 and D498, who were elder compared to others, have weaker responses in LLN compared to other sites (Poon et al., 2021).


Figure 3: Heatmap of quantification of 50 immune mediators produced in different tissue sites of four seropositive donors, stimulated by peptide MPs including MP_S (S), MP_CD4_R (R), MP_CD8_A (A), and MP_CD8_B (B). Figure from Poon et al., 2021.
There was specificity between tissues in the production of mediators stimulated by SARS-CoV-2 peptide pools (Poon et al., 2021). As seen in Figure 4, in the LLN and gut lymph nodes (GLN), the most diverse functional responses were observed with increased levels of mediators, including type 1 pro-inflammatory cytokines and cytolytic mediators, type 2 and type 3 cytokines (Poon et al., 2021). In the lung, there were distinct responses from other sites, including proinflammatory profiles, IL-10, and relatively higher levels of IL-6 and IL-15 (Poon et al., 2021). The bone marrow (BM) and blood have similar responses, including TNF-α, perforin, granzyme B, and IL-10 (Poon et al., 2021). By comparing the milieu of immune mediators among sites, some cytokines are produced in different sites while others are distinct to specific sites (Poon et al., 2021).

Figure 4: Heatmap of immune mediator milieu of different sites. Type 1 proinflammatory cytokines and cytolytic mediators includes IFN-γ, TNF-α, granzyme B, perforin, and GM-CSF. Type 2 cytokines includes IL-5, , IL-9, and IL-13. Type 3 cytokines includes IL-17A, IL-17F, and G-CSF. Proinflammatory profiles in lung includes TNF-α, perforin, granzyme, IL-12, IL-17A, and G-CSF. IFN-γ = interferon-γ; TNF-α = tumor necrosis; GM-CSF = granulocyte-macrophage colony-stimulating factor; IL-5 = interleukin-5; G-CSF = granulocyte colony-stimulating factor. Figure from Poon et al., 2021.
SARS-CoV-2–specific memory B cells and resident phenotypes in tissues
Memory B cells that were specific to SARS-CoV-2 were detected at significantly higher frequencies in all tissues examined in seropositive donors than in seronegative donors as seen in Figure 5A (Poon et al., 2021). Examined tissues include lung, BM, spleen, LLN, and GLN (Poon et al., 2021). IgM+, IgA+ and IgG+ are the isotypes of SARS-CoV-2–specific memory B cells present in the samples with IgG+ being the predominant one as seen in Figure 5B (Poon et al., 2021). The frequencies of these memory B cells were higher in lung and LLNs compared to spleen and GLNs (Poon et al., 2021). Two points worth mentioning is that, for donor D498, few IgM+ in the LNs, and more than 40% were IgM+ in the BM and spleen; for donor D495, no IgA+ in the lung, and about 25% were IgA+ in GLNs (Poon et al., 2021).

Figure 5: A Frequencies of S/RBD-specific memory B cells in tissues, expressed as a percentage of CD19+CD20+ total B cells. There were 4 seropositive donors (n = 4), with n = 4 for lung and LN, n = 3 for spleen, and n = 2 for BM. There were 7 seronegative donors (n = 7), with n = 5 for LN and BM, n = 4 for lung and spleen. B Percentages of each Ig isotypes detected in different sites from four donors. S/RBD = Spike and/or RBD. Figure from Poon et al., 2021.
Memory B cells can persist in lymphoid or non-lymphoid tissues as tissue-resident cells (BRM) and are recognized by CD69 expression (Poon et al., 2021). In tissues from seropositive and seronegative donors, about 50 to 80% of CD69-expressing B cells were detected in the lung and approximately 20 to 40% were detected in LNs (Poon et al., 2021). By contrast, the frequencies of CD69+ SARS-CoV-2 S/RBD–specific memory B cells were much lower in BM and spleen, which fell within the range from 0 to 10% (Poon et al., 2021). This was summarized in Figure 6.

Figure 6: A Percentages of CD69+ SARS-CoV-2 S/RBD–specific memory B cells in different sites. B Flow cytometry plots representation of CD69 expression on S/RBD-specific memory B cells. Figure from Poon et al., 2021.
SARS-CoV-2-specific germinal centre B cells and T cells in lymph nodes
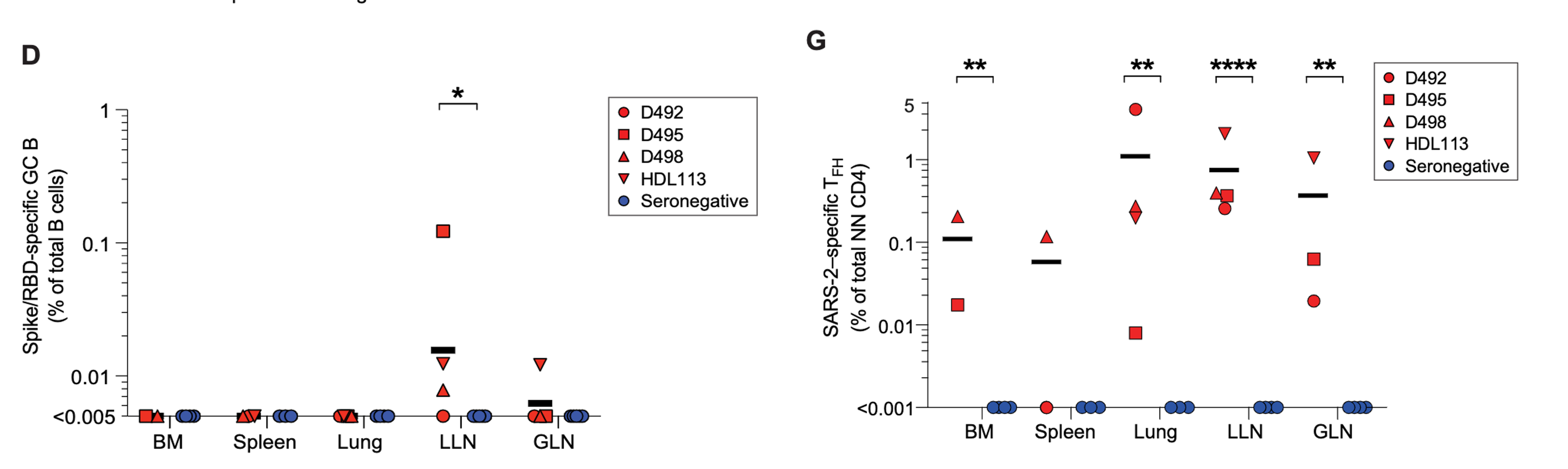
Figure 7: The frequency of S/receptor binding domain-specific germinal centre B cells among total amount of B cells (left). Red represents seropositive donors. The blue represents seronegative donors. The frequency of SARS-CoV-2 specific follicular helper T cells in different tissues among seropositive and seronegative donors (right). Red represents seropositive donors. The blue represents seronegative donors. Figure from Poon et al., 2021.
Activated B cells can receive aid from follicular helper T cells in order to go through somatic hypermutation in germinal centres of lymphoid organs. This will allow them to develop an ameliorated affinity of antibody recognition for pathogens. Despite other studies revolving around the idea, the paper claims that prior to this, there was not a sufficient amount of evidence regarding SARS-CoV-2 antigen-specific germinal centres. More specifically, they were aiming to find S protein specific germinal centre B cells. They identified germinal centre B cells by searching for the co-expression of the transcription factor, Bcl6 and active cellular proliferation marker, Ki67 (Poon et al., 2021). Lung-associated lymph nodes were found to have an increased frequency in germinal centre B cells in seropositive donors than seronegative donors (Poon et al., 2021). Furthermore, SARS-CoV-2 specific germinal centre B cells, which were found using binding to S and receptor binding domain proteins, were also found more frequently in seropositive donors (Poon et al., 2021). Specifically, they were found in lung-associated lymph nodes in 3 out of the 4 seropositive donors (Poon et al., 2021). These cells, however, were not detected in bone marrow, the spleen or directly in the lungs of seropositive donors or any tissues of seronegative donors (Poon et al., 2021). Figure 7 (left) demonstrates the prevalence of B cells across tissues and its presence in lung associated lymph nodes. This demonstrates that SARS-CoV-2 specific germinal centre responses are a result of SARS-CoV-2 infection and are maintained in lung-associated lymph nodes.
A similar result can be seen when investigating follicular helper T cells. In both seronegative and seropositive donors, lymphoid sites such as the spleen, lung-associated lymph nodes and gut associated lymph nodes had very low amounts of follicular helper T cells (Poon et al., 2021). Even lower amounts were seen in bone marrow and most lungs of the same donors (Poon et al., 2021). These cells are very rare among total T cell populations. SARS-CoV-2 specific TFH cells were seen in many sites of the seropositive donors (Poon et al., 2021). They made up 20 to 50% of the SARS-CoV-2 reactive CD4+ T cells in lymph nodes (Poon et al., 2021). The lung, spleen and bone marrow had lower amounts but still a significant frequency of SARS-CoV-2 specific T FH cells (Poon et al., 2021). In one of the seropositive donors, it was found that over 80% of the SARS-CoV-2 specific CD4+ T cells were TFH cells (Poon et al., 2021). Figure 8 demonstrates the prevalence of follicular helper T cells across tissues and the lack of prevalence in seronegative donors. Both points together lead to an implication that there are high germinal centre responses following the infection of SARS-CoV-2 which can be seen in lymphoid areas as well as sites of infection. Some germinal centres are found to be active months following SARS-CoV-2 infection.
Coordinated adaptive immunity across tissues
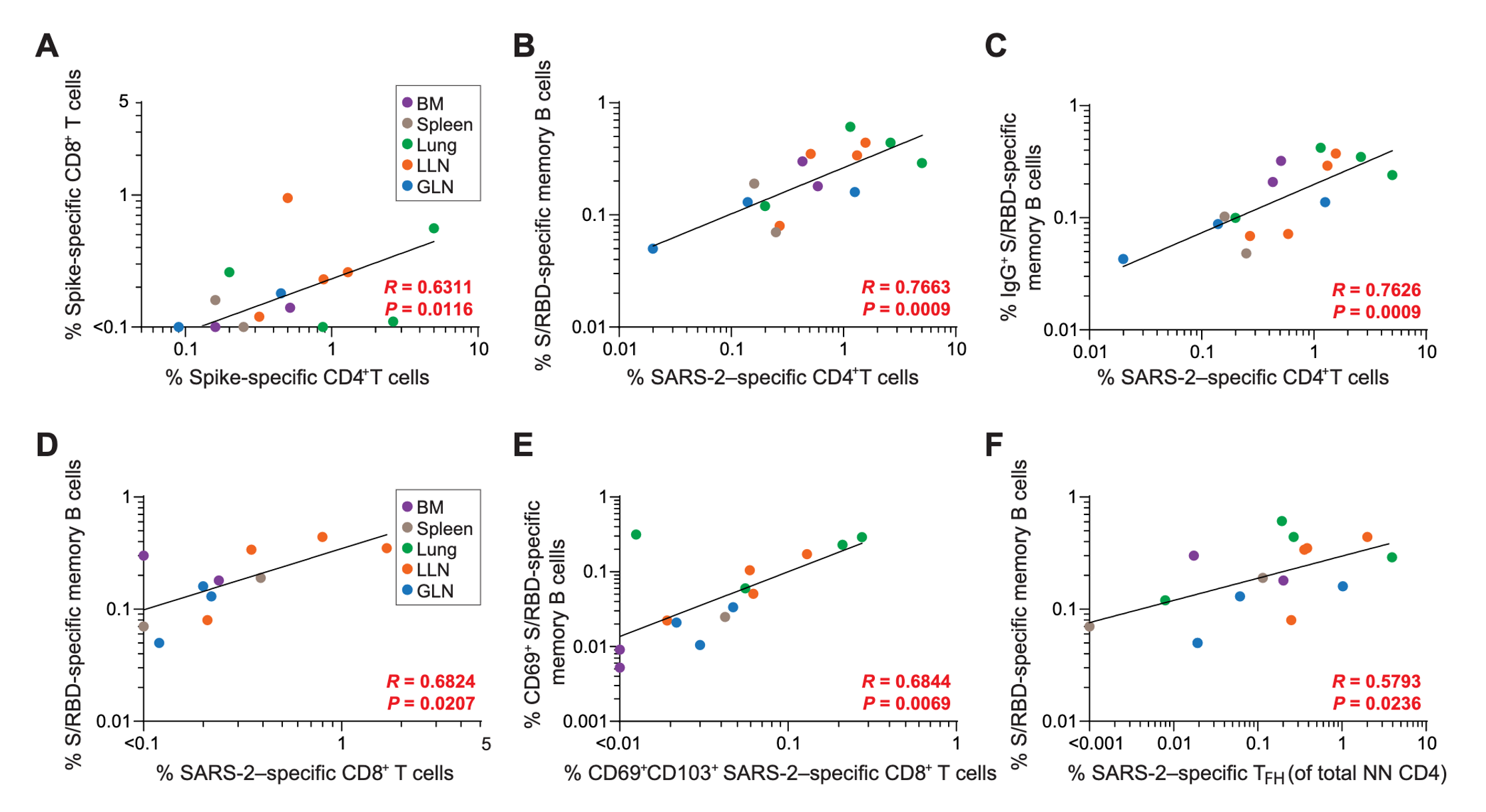
Figure 8: The positive correlations that exist between T cell populations and between T and B cell populations. All correlations listed are SARS-CoV-2 specific lymphocytes found across various tissue types. No negative correlations are shown in figures. Figure from Poon et al., 2021.
An exploratory correlation analysis was conducted in order to identify the associations that existed between the SARS-CoV-2 specific lymphocyte populations across different tissues sites. S-specific CD4+ and CD8+ T cells were seen to be positively correlated across different tissue sites (Poon et al., 2021). Associations between B cells and T cells were also found (Poon et al., 2021). It was found that SARS-CoV-2 specific CD4+ T cell frequencies were positively correlated to the receptor binding domain specific memory B cells in all tissues (Poon et al., 2021). It was also positively correlated with the populations that were IgG+ and IgA+ (Poon et al., 2021). The same positive correlations were also found in T and B resident memory cells in all tissues, as well as follicular helper cells and memory B cells (Poon et al., 2021).
A targeted correlation matrix analysis was also performed to identify associations of SARS-CoV-2 specific lymphocytes within the lung and lung-associated lymph nodes. The follicular helper T cells in the lymph nodes were found to have a significant correlation with memory B cells in the lungs (Poon et al., 2021). CD4+ T cell frequencies in the lungs were also positively correlated to the B cell in germinal centres of the lung associated lymph nodes (Poon et al., 2021). There were also negative correlations that were observed. This includes the relationship between S/receptor binding domain-specific germinal centre B cells in the lymph nodes and the S-specific CD4+ T cells in the lung (Poon et al., 2021). S/RBD specific memory B cell frequencies in the lymph nodes and the S-specific CD8+ T cell frequencies in the lung also has the same relationship (Poon et al., 2021). There are clearly suggested relationships both positively and negatively correlated in terms of immune responses that occur in the lung-associated lymph nodes and the lungs.
Future Directions
The main idea that this study put forward is that memory cells have different responses depending on where they are detected within the body. In the case of SARS-CoV-2, the memory cells in the lungs and lung associated lymph nodes are more specific for SARS-CoV-2 than the memory cells found in other areas of the body. What this could suggest is that future developments of vaccines or treatments could be more specifically targeted to the organs and lymph nodes that they infect. There are already ongoing studies researching an inhalable form of the SARS-CoV-2 vaccine which would strengthen the immune response of the memory cells located in the lungs and lung associated lymph nodes.
Improvements to Experimental Design
The biggest issue with the experimental design of this study was the lack of experimental data. The study obtained samples from people who had to meet the following conditions; they had to be unvaccinated against the SARS-Cov-2 virus, they had to donate live blood samples and organ samples over a long period of time, and they had to be infected with the SARS-CoV-2 virus. These three main conditions makes it difficult for the researchers to find suitable participants in the study. Of the four seropositive donors that they did have however, useful data was generated and general trends were observed, but if the sample size in participants was larger, this data would be even more impactful.
References
Organ procurement organizations | Increasing organ donations. (n.d.). UNOS. Retrieved February 17, 2022, from https://unos.org/transplant/opos-increasing-organ-donation/
Allie, S. R., Bradley, J. E., Mudunuru, U., Schultz, M. D., Graf, B. A., Lund, F. E., & Randall, T. D. (2018). The establishment of resident memory B cells in the lung requires local antigen encounter. Nature Immunology, 20(1), 97–108. https://doi.org/10.1038/s41590-018-0260-6
Dan, J. M., Mateus, J., Kato, Y., Hastie, K. M., Yu, E. D., Faliti, C. E., Grifoni, A., Ramirez, S. I., Haupt, S., Frazier, A., Nakao, C., Rayaprolu, V., Rawlings, S. A., Peters, B., Krammer, F., Simon, V., Saphire, E. O., Smith, D. M., Weiskopf, D., … Crotty, S. (2021). Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. Science, 371(6529). https://doi.org/10.1126/SCIENCE.ABF4063/SUPPL_FILE/PAPV2.PDF
Gray, J. I., Westerhof, L. M., & MacLeod, M. K. L. (2018). The roles of resident, central and effector memory CD4 T‐cells in protective immunity following infection or vaccination. Immunology, 154(4), 574. https://doi.org/10.1111/IMM.12929
Liao, M., Liu, Y., Yuan, J., Wen, Y., Xu, G., Zhao, J., Cheng, L., Li, J., Wang, X., Wang, F., Liu, L., Amit, I., Zhang, S., & Zhang, Z. (2020). Single-cell landscape of bronchoalveolar immune cells in patients with COVID-19. Nature Medicine, 26(6), 842–844. https://doi.org/10.1038/s41591-020-0901-9
Poon, M. M. L., Rybkina, K., Kato, Y., Kubota, M., Matsumoto, R., Bloom, N. I., Zhang, Z., Hastie, K. M., Grifoni, A., Weiskopf, D., Wells, S. B., Ural, B. B., Lam, N., Szabo, P. A., Dogra, P., Lee, Y. S., Gray, J. I., Bradley, M. C., Brusko, M. A., … Farber, D. L. (2021). SARS-CoV-2 infection generates tissue-localized immunological memory in humans. Science Immunology, 6(65), eabl9105. https://doi.org/10.1126/sciimmunol.abl9105
Reiss, S., Baxter, A. E., Cirelli, K. M., Dan, J. M., Morou, A., Daigneault, A., Brassard, N., Silvestri, G., Routy, J.-P., Havenar-Daughton, C., Crotty, S., & Kaufmann, D. E. (2017). Comparative analysis of activation induced marker (AIM) assays for sensitive identification of antigen-specific CD4 T cells. PLoS ONE, 12(10), e0186998. https://doi.org/10.1371/journal.pone.0186998
Teijaro, J. R., Turner, D., Pham, Q., Wherry, E. J., Lefrançois, L., & Farber, D. L. (2011). Tissue-Retentive lung memory CD4 T cells mediate optimal protection to respiratory virus infection. J. Immunol., 187(11), 5510. https://doi.org/10.4049/JIMMUNOL.1102243
Zuo, J., Dowell, A. C., Pearce, H., Verma, K., Long, H. M., Begum, J., Aiano, F., Amin-Chowdhury, Z., Hallis, B., Stapley, L., Borrow, R., Linley, E., Ahmad, S., Parker, B., Horsley, A., Amirthalingam, G., Brown, K., Ramsay, M. E., Ladhani, S., & Moss, P. (2021). Robust SARS-CoV-2-specific T cell immunity is maintained at 6 months following primary infection. Nature Immunology, 22(5), 620–626. https://doi.org/10.1038/s41590-021-00902-8